 STOP MORTALITÉ ÉPILEPSIE
STOPMEP :
STOP MORTALITÉ ÉPILEPSIE
STOPMEP : Modulation de la respiration selon le moment de la journée et implications pour la SUDEP Une étude de K. Joyal B. Kreitlow. Buchanan
Traduction de l’article : The role of sleep state and time of day in modulating breathing in epilepsy : implications for sudden unexpected death in epilepsy. Front Neural Circuits. 2022 Aug 23 ;16:983211. doi : 10.3389/fncir.2022.983211. PMID : 36082111 ; PMCID : PMC9445500.
https://pmc.ncbi.nlm.nih.gov/articles/PMC9445500/
Le rôle de l’état de sommeil et du moment de la journée dans la modulation de la respiration chez les personnes épileptiques : implications pour la mort subite inattendue en cas d’épilepsie
Katelyn G. Joyal (1),(2),(3) - Benjamin L. Kreitlow (1),(2),(3),(4) - Gordon F. Buchanan (1),(2),(3),(4)
(1) Programme interdisciplinaire d’études supérieures en neurosciences, Faculté de médecine Carver, Université de l’Iowa, Iowa City, IA, États-Unis
(2) Département de neurologie, Faculté de médecine Carver, Université de l’Iowa, Iowa City, IA, États-Unis
(3) Institut des neurosciences de l’Iowa, Faculté de médecine Carver, Université de l’Iowa, Iowa City, IA, États-Unis
(4) Programme de formation des chercheurs en médecine, Faculté de médecine Carver, Université de l’Iowa, Iowa City, IA, États-Unis
Résumé :
La mort subite inattendue en épilepsie (SUDEP) est la principale cause de décès chez les patients atteints d’épilepsie réfractaire. Bien que l’étiologie exacte de la SUDEP demeure inconnue, de plus en plus de données suggèrent que les troubles respiratoires jouent un rôle déclencheur dans les cas de décès induits par une crise.
Un trouble de la régulation respiratoire peut survenir chez les patients épileptiques pendant et après les crises, ainsi qu’en dehors de celles-ci. Nombre d’entre eux présentent d’ailleurs des troubles respiratoires du sommeil (TRS), tels que l’apnée obstructive du sommeil (AOS).
La majorité des cas de SUDEP surviennent la nuit, la victime étant retrouvée en position ventrale dans ou près d’un lit.
Étant donné que la respiration varie à la fois en fonction de l’heure de la journée et de l’état de sommeil, il est pertinent d’étudier l’impact supplémentaire des crises nocturnes sur la fonction respiratoire.
Cette revue explore l’état actuel des connaissances sur la relation entre la fonction respiratoire, l’état de sommeil, le moment de la journée, et l’épilepsie. Nous soulignons que le sommeil représente une période particulièrement vulnérable pour les personnes atteintes d’épilepsie et insistons sur le fait que ce sujet mérite d’être approfondi afin de développer des interventions thérapeutiques pour atténuer le risque de SUDEP.
Introduction
L’épilepsie est l’un des troubles neurologiques les plus fréquents. Aux États-Unis, une personne sur 26 développera une épilepsie au cours de sa vie (Kotsopoulos et al.,2002 ; Hesdorffer et al.,2011.)
Malgré sa prévalence, environ 35 % des patients ne parviendront pas à une disparition complète des crises grâce au traitement médical (Kwan et Brodie,2000 ; Chen et al.,2018). Bien que la disponibilité des médicaments antiépileptiques (MAE) ait continué de s’accroître, les patients qui ne répondent pas suffisamment au traitement initial par MAE sont susceptibles de souffrir d’épilepsie pharmacorésistante (Kwan et Brodie,2000). La principale cause de décès chez ces personnes présentant un mauvais contrôle des crises est la mort subite inattendue en épilepsie ou SUDEP (Devinsky et al.,2016). La SUDEP est définie comme « la mort subite, inattendue, avec ou sans témoin, non traumatique et non par noyade, chez les patients épileptiques, avec ou sans preuve de crise, et à l’exclusion de l’état de mal épileptique documenté, pour laquelle l’examen post-mortem ne révèle aucune cause toxicologique ou anatomique de décès » (Nashef et al.,1998).
Bien que, par définition, la SUDEP ne survienne pas nécessairement après une crise d’épilepsie, de solides preuves suggèrent qu’elle en fait un phénomène lié aux crises d’épilepsie, dont les mécanismes agoniques débutent pendant ou immédiatement après une crise (Nashef et al.,1998 ; Nilsson et al.,1999 ; Surges et al.,2009 ; Surges et Sander,2012 ; Bozorgi et Lhatoo,2013). On observe une légère prédominance des cas de SUDEP chez les hommes par rapport aux femmes (Tennis et al.,1995 ; Nilsson et al.,1999 ; Shankar et al.,2013).
Malgré le fardeau considérable que représente la SUDEP, ses mécanismes pathologiques sous-jacents sont mal compris. Cependant, des preuves s’accumulent qui impliquent l’insuffisance respiratoire liée aux crises d’épilepsie comme facteur majeur de ce phénomène mortel (Ryvlin et al.,2013 ; Buchanan et al.,2014 ; Kim et al.,2018 ; Dhaibar et al.,2019).
Dans les cas de SUDEP enregistrés dans les unités de surveillance de l’épilepsie (USE), l’apnée terminale a précédé l’asystolie terminale dans tous les cas (Ryvlin et al.,2013). De plus, il a été constaté que la ventilation mécanique réduisait considérablement la mortalité induite par les crises d’épilepsie, tant chez les patients humains que dans les modèles animaux (Tupal et Faingold,2006 ; Ryvlin et al.,2013 ; Buchanan et al.,2014). Ainsi, des recherches plus approfondies sur les dysfonctionnements respiratoires dans l’épilepsie sont essentielles pour élucider les mécanismes sous-jacents de la SUDEP et pour aider les cliniciens à développer des interventions axées sur la fonction respiratoire.
Une autre observation constante est que les cas de SUDEP surviennent principalement pendant la nuit (Nobili et al.,2011 ; Lamberts et al.,2012 ; Sveinsson et al.,2018). Environ 95 % des cas de SUDEP surviennent au domicile de la victime, la majorité des victimes étant retrouvées dans ou près d’un lit, en position ventrale (Opeskin et Berkovic,2003 ; Zhuo et coll.,2012 ; Ali et al.,2017 ; Sveinsson et al.,2018). Bien que se produisant si près de leur domicile, la grande majorité de ces cas se produisent sans témoins (Lamberts et al.,2012 ; Zhuo et coll.,2012 ; Rugg-Gunn et al.,2016 ; Purnell et al.,2018).
Les patients décédés de SUDEP ont deux fois plus de risques d’avoir des antécédents de crises nocturnes ; la présence de crises nocturnes est donc considérée comme un facteur de risque de SUDEP (Lamberts et al.,2012 ; Shankar et al.,2013 ; Sveinsson et al.,2018 ; Van Der Lende et al.,2018).
Les crises et les décharges épileptiformes surviennent plus fréquemment pendant le sommeil paradoxal (NREM) chez les patients humains et les modèles animaux (Bazil et Walczak,1997 ; Malow et al.,1998). L’état de sommeil peut influencer la fréquence, la gravité et la durée des crises (Bazil et Walczak,1997 ; Ng et Pavlova,2013). Les crises survenant pendant le sommeil ont tendance à être plus longues et sont plus susceptibles d’évoluer en crises tonico-cloniques focales et bilatérales (Bazil et Walczak,1997).
Comme les humains ont tendance à consolider leur sommeil pendant la nuit, de nombreuses études et conclusions sur les facteurs de risque de SUDEP confondent état de sommeil et période nocturne. En réalité, le sommeil et le rythme circadien peuvent modifier indépendamment les processus physiologiques, notamment les fonctions respiratoires et cardiaques (Snyder et al.,1964 ; Browne et al.,1983 ; Spengler et al.,2000 ; Mortola,2004 ; Buchanan,2013). L’influence majeure du sommeil et du rythme circadien sur la respiration en fait un point d’examen essentiel dans le cadre de la physiopathologie de la SUDEP. L’objectif de cette revue est d’examiner les influences distinctes du sommeil et des rythmes circadiens sur la respiration, tant dans un cerveau sain que chez les patients épileptiques (cf figure )
Nous espérons non seulement mettre en évidence les facteurs qui rendent les crises nocturnes plus mortelles, mais aussi mieux différencier les influences de l’état de sommeil et du moment de la journée sur la respiration, afin que les cliniciens puissent développer des stratégies préventives spécifiques pour les dysfonctionnements respiratoires mortels induits par les crises.
Figure 1
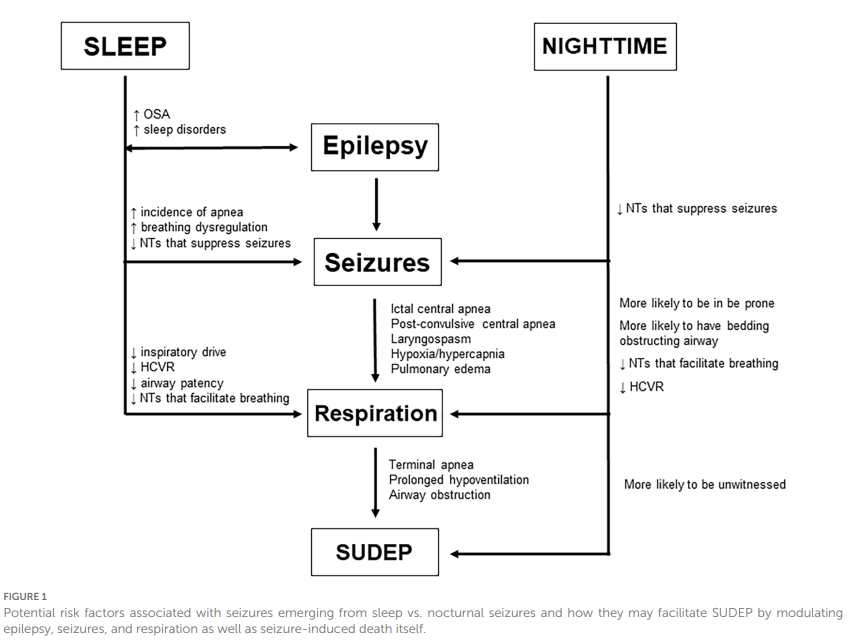
Facteurs de risque potentiels associés aux crises survenant pendant le sommeil par rapport aux crises nocturnes et comment ils peuvent favoriser la SUDEP en modulant l’épilepsie, les crises et la respiration, ainsi que la mort induite par la crise elle-même.
Influence du sommeil sur la respiration
Il est reconnu depuis longtemps que la respiration est régulée en fonction de l’état de sommeil (Snyder et al.,1964 ; Spengler et al.,2000 ; Haxhiu et al.,2003 ; Mortola,2004 ; Malik et al.,2012 ;Buchanan,2013). La stimulation inspiratoire est plus faible pendant le sommeil NREM et minimale pendant le sommeil paradoxal (REM), le volume courant (VC) étant réduit à 73 % de son niveau pendant l’éveil (Douglas et al.,1982a ;Figure 2A
Au cours du sommeil NREM, le nadir de la ventilation minute (VE) survient pendant le stade 3 (N3) du sommeil NREM, probablement en raison de la réduction du volume courant (VC). Il en résulte une concentration de dioxyde de carbone en fin d’expiration (ETCO2) supérieure de 1 à 2 torr à celle de l’état d’éveil (Krieger, 2005). Cette diminution de VC et VE est probablement due à une baisse de la chimiosensibilité lors de l’endormissement (Bulow,1963 ; Douglas et al.,1982b,c). Pendant le sommeil, on observe une diminution de la réponse respiratoire à l’hypercapnie (Reed et Kellogg,1958 ; Birchfield et al.,1959Cherniack,1981 ; Douglas et al.,1982c Berthon-Jones et Sullivan,1984 ;Figure 3A) ainsi que l’hypoxie (Berthon-Jones et Sullivan,1982 ; Douglas et al.,1982b ; Malik et al.,2012). Tout comme la commande inspiratoire, la diminution de la commande respiratoire induite par l’hypoxie est encore plus importante pendant le sommeil paradoxal (REM) que pendant le sommeil lent (NREM) (Berthon-Jones et Sullivan,1984 ; Malik et al.,2012). Il existe des différences liées au sexe dans la réponse à l’hypercapnie : les hommes présentent une diminution de 50 % de la réponse ventilatoire hypercapnique (RVH) par rapport à l’état de veille, tandis que les femmes présentent une RVH réduite à l’état de veille par rapport aux hommes, mais des réductions moins marquées de la réponse pendant le sommeil (Berthon-Jones et Sullivan,1984). Il a été démontré que la progestérone stimule la respiration pendant le sommeil, notamment en augmentant les réponses respiratoires hypoxiques et hypercapniques (Javaheri et Guerra,1990 ; Saaresranta et al.,1999).
La progestérone oscille selon un rythme circadien, avec un pic aux alentours de minuit (Junkermann et al.,1982 ; Gharib et al.,2018). Aucune différence liée au sexe n’a été identifiée dans les réponses respiratoires à l’hypoxie (Malik et al.,2012).
Figure 2

Effets circadiens et dépendants de l’état de sommeil sur la ventilation.(UN) Tracés sur 72 h de la ventilation minute moyenne (en haut), du volume courant (au milieu) et de la fréquence respiratoire (en bas) chez des rats mâles adultes maintenus sous un cycle lumière/obscurité de 12 h/12 h et recevant de l’air ambiant (21 % d’O₂).2, solde N2Les barres horizontales pleines en bas indiquent les périodes où les lumières étaient éteintes.(B) Enregistrement sur 24 heures de la ventilation minute moyenne chez le rat pendant l’éveil, le sommeil lent (NREM) et le sommeil paradoxal (REM), comme indiqué. Tous les animaux étaient maintenus dans un cycle lumière/obscurité de 12 h/12 h.(UN) Redessiné avec la permission de Seifert et Mortola (2002).(B) Redessiné avec l’autorisation de Stephenson et al. (2001).
Figure 3
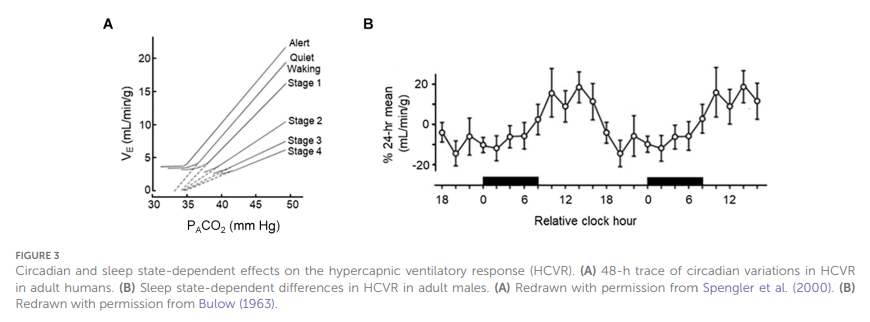
Effets circadiens et liés à l’état de sommeil sur la réponse ventilatoire à l’hypercapnie (HCVR). (A) Tracé sur 48 heures des variations circadiennes de la HCVR chez l’adulte. (B) Différences de la HCVR liées à l’état de sommeil chez les hommes adultes. (A) Redessiné avec l’autorisation de Spengler et al. (2000). (B) Redessiné avec l’autorisation de Bulow (1963).
La respiration pendant le sommeil NREM présente un schéma plus régulier que la respiration pendant l’éveil, sans modifier la fréquence respiratoire moyenne (Malik et al.,2012). À l’inverse, pendant le sommeil paradoxal, les schémas respiratoires présentent une plus grande variabilité, notamment une fréquence accrue, une régularité diminuée et de brèves périodes d’apnée centrale (Aserinsky et Kleitman,1953Cherniack,1981 ; Malik et al.,2012). Certaines données suggèrent que cette respiration irrégulière est une réponse à des entrées corticales qui reflètent le contenu du rêve de l’individu (Oudiette et al.,2018).
La respiration périodique, caractérisée par des séries de respirations séparées par des intervalles d’apnée centrale ou de quasi-apnée, survient parfois aussi pendant le sommeil. Bien qu’on ait longtemps pensé qu’elle résultait d’une affection neurologique ou cardiovasculaire grave, on a maintenant constaté qu’elle pouvait se produire chez des individus sains, notamment en cas d’hypoxie (Berssenbrugge et al.,1983 , Cherniack,1999 ; Ainslie et al.,2013). Lors des phases de respiration périodique, les variations cycliques de la ventilation ainsi que des pressions partielles de dioxyde de carbone (CO2) et d’oxygène (O2) peuvent déclencher des oscillations de la fréquence cardiaque, de la pression artérielle, de l’activité du système nerveux autonome et de la résistance des voies aériennes supérieures. Ceci peut créer une boucle de rétroaction par laquelle ces oscillations, à leur tour, affectent la ventilation et augmentent la durée et la symétrie de ces cycles respiratoires périodiques (Cherniack,1999). Les mâles ont tendance à présenter une respiration périodique en réponse à l’hypoxie plus fréquemment que les femelles (Pramsohler et al., 1997).al.,2019). Les schémas respiratoires dépendent fortement du complexe pré-Bötzinger (pré-BötC ; Smith et al.,1991 ; Buchanan,2013 ; Del Negro et al.,2018 ; Muñoz-Ortiz et al.,2019).
Après l’ablation bilatérale des neurones exprimant les récepteurs de la neurokinine-1 (NK1R) chez des rats adultes, on observait une perturbation progressive et irréversible de la stabilité respiratoire, qui survenait initialement uniquement pendant le sommeil, mais finissait par entraîner une respiration ataxique également à l’état de veille (McKay et Feldman,2008). En revanche,après l’ablation unilatérale de ces neurones pré-BötC exprimant NK1R, on observait une perturbation du rythme respiratoire et une augmentation de la fréquence des apnées et hypopnées centrales du sommeil uniquement pendant le sommeil, en particulier pendant le sommeil paradoxal, qui ne se développaient jamais à l’état de veille (McKay et Feldman,2008).
Pendant le sommeil, la perméabilité des voies aériennes supérieures diminue et la résistance respiratoire augmente. Ceci est dû à une réduction préférentielle du tonus des muscles laryngés et pharyngés qui contribuent au maintien de la structure des voies aériennes supérieures (Cherniack,1981 ; Haxhiu et al.,1987 ; Buchanan,2013 ; Kubin,2016). Cette perméabilité réduite peut s’avérer particulièrement problématique pendant le sommeil paradoxal, lorsque la respiration est particulièrement instable (Cherniack,1981). Le tonus des voies aériennes supérieures est contrôlé par les afférences des motoneurones trigéminaux (V), faciaux (VII) et hypoglosses (XII) (Buchanan,2013). Le muscle génioglosse, innervé par les motoneurones hypoglosses, est le plus grand et le plus étudié des muscles dilatateurs des voies respiratoires. Il a été suggéré que la diminution des afférences sérotoninergiques et noradrénergiques aux motoneurones hypoglosses pendant le sommeil paradoxal provoque l’atonie du génioglosse (Fenik et al.,2005). Le génioglosse et les autres muscles des voies aériennes supérieures nécessitent une activation inspiratoire à la fois tonique et phasique afin de se protéger contre l’affaissement (Kubin,2016). Lorsque le tonus de ces muscles dilatateurs des voies respiratoires ne peut plus s’opposer à la pression inspiratoire négative, il en résulte une apnée obstructive du sommeil (AOS), caractérisée par des épisodes récurrents d’hypopnées et d’apnées (Remmers et al.,1978 ; Kubin,2016). Bien que ces apnées obstructives ne surviennent que pendant le sommeil, des apnées du sommeil fréquentes et une hypoventilation peuvent entraîner des anomalies respiratoires à l’état d’éveil (Simonds,1994).
Des modifications de plusieurs mécanismes respiratoires non centraux sont également associées à l’endormissement. Pendant le sommeil NREM, l’activité des muscles intercostaux est accrue par rapport à l’état de veille (Malik et al.,2012). Cela peut indiquer une contribution accrue de la paroi thoracique à la respiration afin de compenser la diminution de la commande inspiratoire centrale. Pendant le sommeil paradoxal, on observe une perte d’activité tonique au niveau des muscles intercostaux et du diaphragme (Tusiewicz et al.,1977 ; Bryan et Muller,1980 ; Malik et al.,2012). La compliance de la paroi thoracique augmente également pendant cette période et, associée à une diminution du tonus intercostal, peut provoquer un affaissement paradoxal de la cage thoracique lors de l’inspiration (Malik et al.,2012). Enfin, le réflexe d’étirement des récepteurs pulmonaires et le réflexe des récepteurs d’irritation sont supprimés pendant le sommeil ; ainsi, la toux en réponse à l’apnée ne survient qu’après un éveil (Douglas,2000). En résumé, le sommeil est une période où de nombreux aspects de la respiration sont inhibés, ce qui en fait une période particulièrement vulnérable à d’autres agressions du système respiratoire.
Influences circadiennes sur la respiration
Les premières études sur la variabilité diurne de la physiologie respiratoire des mammifères (rats adultes) ont révélé des différences diurnes dans la respiration ; cependant, l’effet était limité à la production de CO2 et au débit d’air inspiratoire moyen (Peever et Stephenson,1997). Dans des conditions d’hypercapnie, la fréquence respiratoire et la ventilation semblaient également dépendre du moment de la journée. Malheureusement, ces études ne portaient que sur deux points temporels, ce qui limitait la résolution d’un rythme circadien, lequel pouvait être masqué par des variations ultradiennes de fréquence plus élevée de la respiration (Stupfel et Pletan,1983 ; Stupfel et al.,1985).
La première preuve claire que la fonction respiratoire présentait des oscillations quotidiennes provient de Seifert et al. (2000). Des rats adultes ont été placés dans des chambres barométriques de 10 L avec un contrôle précis des flux de gaz entrant et sortant, permettant la mesure de leur physiologie respiratoire sur plusieurs jours. Des différences nycthémérales nettes ont été observées au niveau de la fréquence respiratoire, du volume courant (VC) et de la ventilation (VE). Les valeurs les plus élevées ont été observées pendant la phase nocturne, coïncidant avec une température et une activité élevées. Ces résultats ont été approfondis dans une étude ultérieure, démontrant que la consommation d’O2 (une mesure de la consommation d’oxygène) était influencée par la consommation d’oxygène.L’activité métabolique), le temps inspiratoire et le temps expiratoire ont également varié au cours de la journée (Seifert et Mortola, 2002 ;Figure 2B). Il est intéressant de noter que la prise en compte du niveau d’activité n’a pas permis d’éliminer l’influence du moment de la journée sur la ventilation (VE), le volume courant (VC), la fréquence respiratoire ou la consommation d’oxygène. Les auteurs concluent que la variabilité quotidienne observée au niveau de la respiration (et plus particulièrement de la ventilation) est probablement due à d’autres variables physiologiques fluctuant au cours de la journée, telles que la température et la consommation d’oxygène.
En utilisant des méthodes similaires à celles de Seifert et al.,2000, une surveillance respiratoire à long terme chez les primates non humains a également été réalisée (Iizuka et al.,2010). Une pléthysmographie corporelle totale a été réalisée chez 11 macaques cynomolgus mâles non anesthésiés et non contraints. À l’instar des résultats obtenus chez les rats adultes, plusieurs paramètres respiratoires, dont la fréquence respiratoire, le volume courant (VC) et la ventilation (VE), ont varié en fonction du moment de la journée. Cependant, les enregistrements n’ont été effectués que toutes les heures, et il n’est pas certain que l’état de sommeil ait été pris en compte.
Une variabilité diurne de plusieurs paramètres respiratoires a également été démontrée chez l’homme (Spengler et Shea,2000 ; Spengler et al.,2000). Dans un environnement de laboratoire rigoureusement contrôlé, incluant la suppression des repères temporels externes (à l’exception de l’éclairage), une température ambiante constante, un apport alimentaire contrôlé et des horaires de sommeil strictement réglementés, les variations temporelles de la température rectale et du cortisol plasmatique ont été utilisées comme marqueurs circadiens endogènes. L’ETCO2, la consommation d’O2 et la production de CO2 ont toutes présenté des oscillations au cours des 24 heures, avec des niveaux maximaux le matin. De manière intéressante, une variabilité diurne de l’amplitude de la réponse ventilatoire à l’hypoxie (HCVR) a également été observée, un résultat précédemment démontré chez le rat adulte éveillé (Peever et Stephenson, 2011).1997 ;Figure 3B), ce qui suggère que la chimiosensibilité influençant la respiration pourrait également être soumise à une régulation circadienne. La sensibilité à une stimulation hypoxique isocapnique présente certaines indications de dépendance à l’heure de la journée ; cependant, l’effet est beaucoup moins prononcé (Siekierka et al.,2007).
Bien que les études décrites ci-dessus chez le rat, le singe et l’homme démontrent une variation temporelle de la physiologie respiratoire, elles n’ont pas contrôlé l’effet rythmique de la lumière. Par conséquent, il est impossible de conclure si cette variabilité est due à l’effet d’un signal temporel externe ou à celui d’un rythme circadien endogène. La première étude sur la dépendance de la physiologie respiratoire à l’heure de la journée, prenant en compte l’influence de la lumière, a été réalisée chez la couleuvre rayée (Hicks et Riedesel, 1997).1983). Les animaux étaient maintenus soit dans un cycle lumière-obscurité de 14 h/10 h, soit dans l’obscurité constante. Dans ces conditions, il a été constaté que la variabilité diurne de la consommation d’oxygène, de la fréquence respiratoire, du volume courant (VC) et de la ventilation (VE) persistait même dans l’obscurité constante, suggérant une régulation endogène de ces paramètres respiratoires.
Il a été démontré que cette variation de la respiration en fonction du moment de la journée est endogène et circadienne chez la souris, et qu’elle est médiée par le stimulateur circadien central de l’organisme, le noyau suprachiasmatique (Purnell et Buchanan,2020). Des souris C57BL/6J ont été maintenues dans des environnements à cycle lumière-obscurité de 12 h/12 h ou dans l’obscurité constante. Des roues d’exercice ont été utilisées pour évaluer le début de la phase active et déterminer les rythmes locomoteurs individuels en activité libre. L’état de sommeil influençant la respiration (comme décrit en détail précédemment), les mesures respiratoires ont été effectuées uniquement lorsque les animaux étaient éveillés. Une variabilité diurne de la fréquence respiratoire et de l’intensité ventilatoire (IV), mais pas du volume courant (VC), a été observée. La fréquence et l’IV étaient maximales pendant la phase obscure de la journée. Ce rythme diurne s’est avéré circadien, car ces deux rythmes persistaient même lorsque les animaux étaient maintenus dans l’obscurité constante. La lésion électrolytique du noyau suprachiasmatique a éliminé ces rythmes respiratoires, suggérant que la variation circadienne de la respiration est contrôlée par ce noyau.
Bien que le noyau suprachiasmatique soit fréquemment considéré comme l’oscillateur circadien principal, des noyaux situés en dehors du noyau suprachiasmatique et des tissus périphériques peuvent contenir des horloges circadiennes autonomes (Mohawk et al.,2012). De telles horloges périphériques ont été décrites dans les neurones du tronc cérébral et de la moelle épinière impliqués dans la coordination et la production des signaux nécessaires au maintien d’une respiration normale. En mesurant des transcrits du gène de l’horloge moléculaire, tels que…Clock ,Bmal1, et Per, 1/2, des chercheurs ont identifié des rythmes robustes des gènes de l’horloge moléculaire dans le noyau du tractussolitarius (Kaneko et al.,2009 ; Chrobok et al.,2020), noyau moteur phrénique (Kelly et al.,2020), et les tissus laryngés, trachéaux, bronchiques et pulmonaires à l’intérieur des voies respiratoires (Bando et al.,2007). Bando et al. (2007) a démontré que l’horloge périphérique des tissus des voies respiratoires pouvait devenir arythmique suite à une lésion électrolytique du noyau suprachiasmatique. De même, De même, les souris knock-out pour les gènes Clock 1/Clock2, génétiquement arythmiques, ne montraient pas de rythmicité périphérique de l’expression des gènes d’horloge dans les tissus des voies respiratoires.
En conclusion, la phase circadienne exerce une influence importante sur la respiration, indépendamment de l’état de vigilance.
Effets des crises d’épilepsie sur la respiration
Certains patients épileptiques présentent des anomalies respiratoires au repos qui peuvent s’aggraver lors d’une crise. Sainju et al. ont constaté une réponse ventilatoire hypercapnique atténuée chez un sous-groupe de patients épileptiques, les exposant à un risque accru d’hypoventilation péri-ictale (Sainju et al.,2019). Les patients atteints du syndrome de Dravet (DS) présentent également une diminution de la réponse ventilatoire au CO2 (Kim et al.,2018). Plusieurs modèles animaux d’épilepsie présentent une dysrégulation respiratoire, même en l’absence de crise. Le modèle murin Kcna1-null présente un dysfonctionnement respiratoire progressif avec l’âge (Simeone et al.,2018). Tout comme leurs homologues humains, le modèle murin knock-in humain Scn1aR1407X/+ du syndrome de Down présente une réponse ventilatoire réduite au CO₂, ainsi qu’une hypoventilation et une apnée de base (Kuo et al.,2019). Une perte similaire de la réponse ventilatoire hypercapnique a été observée chez des animaux ayant subi une stimulation amygdalienne (Totola et al.,2019). Hajek et Buchanan (2016).
Des études ont montré que les souris présentant une variabilité accrue de leur fréquence respiratoire basale sont plus susceptibles de mourir après une crise d’épilepsie induite par électrochoc maximal (MES). Ces résultats confortent l’hypothèse selon laquelle un dysfonctionnement respiratoire intercritique pourrait servir de biomarqueur chez les personnes présentant un risque plus élevé de mort subite inattendue en épilepsie (SUDEP).
Les crises elles-mêmes peuvent provoquer des altérations profondes de la respiration, notamment la toux, l’apnée, l’hyperventilation, les spasmes bronchiques, l’augmentation de la pression vasculaire pulmonaire, le laryngospasme et l’œdème pulmonaire (Bayne et Simon,1981 ; Kennedy et al.,2015 ; Nakase et al.,2016 ; Rugg-Gunn et al.,2016).
Les crises semblent provoquer des degrés variables de dysrégulation respiratoire en fonction du type et de l’origine de la crise (Bateman et al.,2008 ; Blum,2009). Une durée plus longue des crises est associée à un degré de dysfonctionnement plus important, notamment en ce qui concerne l’hypercapnie, la pression pulmonaire et l’œdème pulmonaire (Bayne et Simon,1981 ; Bateman et al.,2008 ; Seyal et al.,2010 ; Kennedy et al.,2015).
L’hypoventilation pendant une crise peut survenir en raison d’une obstruction des voies respiratoires ou d’une dérégulation des centres respiratoires du cerveau et entraîne généralement une hypercapnie et une hypoxémie (Rugg-Gunn et al.,2016).Les patients atteints du syndrome de Dravet présentent notamment une hypoventilation péri-ictale, qui précède l’apparition de la bradycardie (Kim et al.,2018). L’hypoventilation peut entraîner une insuffisance cardiaque secondaire, notamment lors de crises convulsives où la saturation en oxygène (SaO2) chute en dessous de 90 % (Seyal et al.,2011). L’apnée centrale est une cause possible d’hypoventilation ictale. L’apnée centrale ictale (ACI) est relativement fréquente lors des crises d’épilepsie, en particulier celles impliquant les deux hémisphères cérébraux (Nashef et al.,1996 ; Rugg-Gunn et al.,2016). L’ACI survient exclusivement dans l’épilepsie focale, apparaissant lors de 33 à 50 % des crises focales (Lacuey et al.,2018 ; Vilella et al.,2019 ; Tio et al.,2020). L’ACI peut précéder l’activité épileptique électrographique ainsi que le début clinique de la crise jusqu’à 7 à 10 s (Nishimura et al.,2015 ; Tio et al.,2020). Ces apnées ont tendance à être brèves et n’ont pas d’impact significatif sur la saturation en O2 (Bateman et al.,2008). Une analyse multivariée a indiqué que la propagation controlatérale de la crise et sa durée contribuent mutuellement à l’augmentation de l’ETCO2 qui suit l’ACI (Seyal et al.,2010). Plusieurs modèles animaux d’épilepsie et de SUDEP présentent une ACI, notamment les souris Scn1aR1407X/+, chez lesquelles la ventilation mécanique peut prévenir l’arrêt respiratoire fatal induit par une crise convulsive (Kim et al.,2018). De plus, un modèle d’état de mal épileptique induit chez le mouton présente une ACI et une hypoventilation chez 100 % des animaux, certains entraînant la mort (Johnston et al.,1997). L’apnée centrale post-convulsive (ACPC), en revanche, survient aussi bien dans les épilepsies focales que généralisées, ce qui suggère une physiopathologie distincte de celle de l’ACI (Vilella et al.,2019). L’ACPC est moins fréquente queL’ACI (cause cérébrale aiguë) ne survient que dans 18 % des crises généralisées. Cependant, l’ACPC (cause cérébrale postérieure) peut être beaucoup plus dangereuse que l’ACI. L’ACPC est associée à un temps de récupération plus long après une hypoxémie et est considérée par certains comme un biomarqueur de la SUDEP (Jin et al.,2017 ; Vilella et al.,2019).
Les crises convulsives peuvent altérer la capacité d’une personne à s’autoréanimer après une apnée centrale. L’autoréanimation est un phénomène cardiorespiratoire protecteur spontané qui favorise le rétablissement d’une respiration et d’un rythme cardiaque normaux après une apnée primaire en déclenchant un réflexe de suffocation (Adolph,1969 ; Guntheroth et Kawabori,1975). L’échec de l’autoréanimation a été documenté dans des cas de décès de nourrissons qui ont finalement été classés comme syndrome de mort subite du nourrisson (SMSN ; Meny et al.,1994 ; Sridhar et al.,2003). Il existe de nombreux parallèles entre la mort subite du nourrisson (MSN) et la mort subite inattendue en épilepsie (SUDEP), notamment une autopsie normale, la position ventrale, une prédominance nocturne, un mécanisme respiratoire prédit et des signes de dysfonctionnement du système sérotoninergique (Richerson et Buchanan,2011 ; Buchanan,2019).
L’apnée obstructive, ou laryngospasme, est un autre phénomène associé aux crises d’épilepsie qui peut entraîner la mort (Stewart,2018). Les souris DBA/2, qui présentent des crises audiogènes mortelles, ont un taux de mortalité significativement réduit après les crises suite à l’implantation d’un tube en T trachéal comme voie respiratoire de substitution (Irizarry et al.,2020). Il a été démontré que des crises induites via l’acide kaïnique chez les rats provoque une fermeture glottique partielle ou complète et la mort subséquente (Nakase et al.,2016 ; Budde et al.,2018 ; Jefferys et al.,2019). Il a été postulé que l’apnée obstructive fatale est une conséquence de spasmes bronchiques ou d’une hypotonie des muscles impliqués dans la respiration (Stöllberger et Finsterer,2004). Nakase et al. ont suggéré que le laryngospasme ictal est causé par la propagation d’une crise d’épilepsie via des régions motrices médullaires autonomes des branches laryngées du nerf vague (Nakase et al.,2016).
La propagation de la dépolarisation (PD) pourrait être l’un des mécanismes sous-jacents à l’insuffisance cardiorespiratoire dans la SUDEP. Chez les souris mutantes Cacna1a S218L porteuses d’une mutation à gain de fonction du canal calcique voltage-dépendant Cav2.1, une SD du tronc cérébral survient lors de toutes les crises fatales spontanées, ainsi que dans un sous-ensemble de crises non fatales (Jansen et al.,2019). De plus, la PD liée aux crises dans la moelle ventrolatérale est corrélée à l’incidence de la suppression respiratoire (Jansen et al.,2019). Les crises d’épilepsie induites chimiquement chez les souris mutantes Kcna1 et Scn1a provoquent une vague de PD dans la moelle dorsale, ce qui peut temporairement réduire au silence les cellules qui serviraient à réoxygéner le cerveau après une crise (Aiba et Noebels,2015).Ce blocage dépolarisant peut provoquer une boucle de rétroaction positive dans laquelle le cerveau ne peut pas se réoxygéner après une crise d’épilepsie au cours de laquelle la saturation en oxygène a chuté de façon spectaculaire, ce qui peut potentiellement conduire à un arrêt cardiorespiratoire complet (Aiba et Noebels,2015).
De nombreux autres mécanismes potentiels sous-jacents au dysfonctionnement et à l’insuffisance respiratoire ictale ont été proposés. Une hypothèse majeure est que les crises activent des projections sous-corticales inhibitrices vers les centres respiratoires du tronc cérébral (Dlouhy et al.,2015).Lacuey et al.,2017).Il a également été constaté que l’apnée centrale survient chez les patients humains lorsque les crises se propagent à l’amygdale (Dlouhy et al.,2015 ; Rhone et al.,2020).De même, la stimulation de l’amygdale ainsi que de l’hippocampe produit une apnée centrale dont les patients ne sont pas conscients (Dlouhy et al.,2015).Lacuey et al.,2017 ; Nobis et al.,2018), et ils sont capables de déclencher volontairement une inspiration lorsqu’ils y sont invités (Dlouhy et al.,2015).
Des études complémentaires ont révélé que la stimulation de l’amygdale basale en particulier (y compris les noyaux basomédial et basolatéral) était particulièrement susceptible de provoquer des apnées, tandis que la stimulation de régions plus latérales produisait moins d’apnées (Rhone et al.,2020).Chez les souris DBA/1, des lésions unilatérales de l’amygdale suffisaient à supprimer l’arrêt respiratoire induit par les crises (S-IRA) (Marincovich et al.,2021).Cela suggère que l’apnée est due à la perte de la commande ventilatoire involontaire plutôt qu’à un problème au niveau des voies motrices respiratoires ou de la musculature.
Malgré l’implication des dysfonctionnements respiratoires et cardiaques dans la SUDEP, des données récentes suggèrent que l’insuffisance respiratoire précède l’insuffisance cardiaque lors des décès induits par une crise d’épilepsie. En 2013, une étude multicentrique sur la mortalité dans les unités de surveillance (MORTEMUS) portant sur les cas de SUDEP dans ces unités a révélé que tous les cas enregistrés présentaient un arrêt respiratoire terminal avant l’asystolie terminale (Ryvlin et al.,2013).
Des résultats similaires ont été obtenus dans le modèle de souris Kcna1-nul (Dhaibar et al.,2019) et dans un modèle MES (Buchanan et al.,2014).
Un autre indicateur du rôle crucial de l’insuffisance respiratoire dans la SUDEP est que la ventilation mécanique, si elle est administrée immédiatement, peut réduire considérablement la mortalité chez les patients humains et dans les modèles animaux (Tupal et Faingold,2006 ; Ryvlin et al.,2013 ; Buchanan et al.,2014). De même, l’oxygénation avant l’induction de la crise peut prévenir les crises audiogènes fatales chez plusieurs souches de souris audiogènes, sans incidence sur la fréquence ou la gravité des crises (Venit et al.,2004).En résumé, les crises d’épilepsie provoquent des altérations profondes de la respiration qui peuvent contribuer directement à la mort induite par les crises.
Effets des crises d’épilepsie sur le sommeil et le rythme circadien, et leur impact sur la respiration
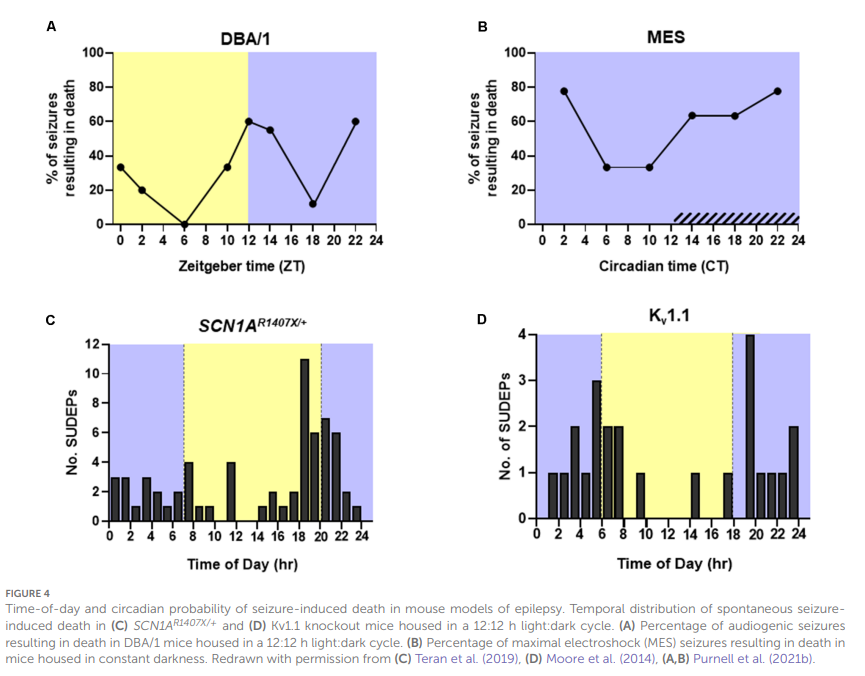
Environ 10 à 15 % des patients épileptiques présentent des crises uniquement ou principalement pendant le sommeil (Grigg-Damberger et Foldvary-Schaefer, 2021).Les crises survenant pendant le sommeil ont tendance à être plus longues et sont plus susceptibles d’évoluer en crises tonico-cloniques focales et bilatérales (Bazil et Walczak,1997).Comme mentionné précédemment, les crises convulsives plus longues sont associées à un degré accru de dysfonctionnement respiratoire (Bayne et Simon,1981 ; Bateman et al.,2008 ; Seyal et al.,2010 ; Kennedy et al., 2015).Les crises survenant pendant le sommeil sont également plus susceptibles d’être associées à la présence d’une suppression généralisée post-critique de l’EEG (PGES) et à une désaturation en oxygène plus importante (Latreille et al.,2017). Une étude clinique menée auprès de 20 patients épileptiques a révélé que 44 % des crises nocturnes sont associées à une ACI, et bien que la différence n’ait pas atteint le seuil de signification statistique, une plus petite proportion de crises survenant à l’éveil était accompagnée d’ACI (28 % ; Latreille et al.,2017).Les crises d’épilepsie induites par MES chez la souris pendant le sommeil NREM sont également associées à un dysfonctionnement respiratoire plus important que celles induites pendant l’éveil (Hajek et Buchanan,2016).En tenant compte du moment de la journée, les crises d’épilepsie induites par stimulation électrique mésenchymateuse (MES) pendant la journée, phase d’inactivité du rongeur, ont entraîné une suppression respiratoire et électroencéphalographique post-critique plus importante que celles induites pendant la nuit. Cet effet était encore plus marqué lorsque les crises étaient induites pendant le sommeil paradoxal (NREM) (Purnell et al.,2017). Lorsque des souris DBA/1, modèle de crises audiogènes, ont été exposées à un stimulus audiogène pendant la journée, les crises qui en ont résulté ont entraîné la mort dans 21,7 % des essais. À l’inverse, les crises induites pendant la nuit ont entraîné la mort par crise dans 46,7 % des essais (Purnell et al.,2021b ;Figure 4A.
La même étude a utilisé des souris vivant dans l’obscurité constante pour évaluer l’influence du rythme circadien sur la mort induite par les crises d’épilepsie dans le modèle murin MES. Ils ont constaté que pendant la nuit subjective, il y avait une diminution de la ventilation post-critique et une augmentation de la probabilité de mort induite par les crises d’épilepsie sans altérer la gravité des crises (Purnell et al.,2021b ;Figure 4B). Les souris Kv1.1 knockout (KO) et SCN1A R1407X/+, qui présentent une dysrégulation respiratoire progressive (Kim et al.,2018 ; Kuo et al.,2019 ; Iyer et al.,2020), subissent également plus fréquemment des décès par crise d’épilepsie pendant la nuit (Figures 4C,D).
Figure 4
Comme les humains dorment généralement la nuit, les crises nocturnes passent souvent inaperçues (Lamberts et al.,2012 ; Zhuo et coll.,2012 ; Rugg-Gunn et al.,2016 ; Purnell et al.,2018). Lamberts et al. (2012) ont rapporté que 86 % des cas de SUDEP surviennent sans témoin. On suppose que le fait d’être seul lors d’une crise nocturne peut comporter un risque encore plus important que la gravité du dysfonctionnement respiratoire lié au sommeil ou la durée de la PGES (Peng et al.,2017 ; Sveinsson et al.,2020). La présence d’une personne capable d’intervenir et d’administrer des mesures de réanimation vitales peut faire la différence entre un cas de SUDEP imminent et un SUDEP avéré (Nashef et al.,1998 ; Langan et al.,2005 ; Lamberts et al.,2012). L’augmentation de la surveillance nocturne par l’utilisation de dispositifs de surveillance, de contrôles réguliers ou la présence d’une autre personne dormant dans la même pièce est associée à une diminution du risque de SUDEP (Langan et al.,2005 ; Ryvlin et al.,2006 ; Harden et al.,2017La majorité des victimes de SUDEP sont retrouvées allongées sur le ventre dans ou près d’un lit (Opeskin et Berkovic,2003 ; Sowers et al.,2013 ; Ali et al.,2017 ; Sveinsson et al.,2018). De nombreuses crises généralisées sont suivies d’une période de PGES où le patient est plus susceptible d’être immobile, inconscient et de nécessiter des mesures de réanimation (Semmelroch et al.,2012 ; Kuo et al.,2016). Si un patient ne réagit pas après une crise convulsive qui le met en décubitus ventral, son nez et sa bouche peuvent être obstrués par la literie. Ceci peut entraîner une occlusion des voies aériennes supérieures ou une asphyxie par contact avec la surface sur laquelle le patient est positionné. En dehors d’une occlusion totale des voies aériennes, le fait de terminer une crise convulsive en position ventrale sur la literie peut altérer la respiration post-critique en augmentant la résistance inspiratoire et en provoquant la réinhalation d’air emprisonné (Kemp et al.,1994 ; Tao et al.,2010,2015 ; Rugg-Gunn et al.,2016). Cela provoquerait une augmentation aiguë du CO2 dans le sang, pouvant entraîner une acidose sévère, ce qui potentialiserait l’immobilité post-critique et prolongerait davantage le dysfonctionnement respiratoire jusqu’à l’apparition d’une apnée terminale et d’une asystolie (Peng et al.,2017 ; Purnell et al.,2018).
Gènes d’horloge
Bien que les crises d’épilepsie soient souvent considérées comme un phénomène imprévisible, les patients présentent souvent un rythme circadien d’apparition des crises. Une étude récente menée auprès de patients porteurs de neuro-stimulateurs réactifs a montré que près de 90 % des patients atteints d’épilepsie focale présentaient un rythme circadien d’apparition des crises (Leguia et al.,2021).Il est intéressant de noter que le risque circadien de déclenchement des crises d’épilepsie pourrait être regroupé en cinq moments généraux de la journée, les crises étant plus susceptibles de survenir le matin, en milieu d’après-midi, le soir, en début de nuit ou tard dans la nuit.
L’influence circadienne sur les crises d’épilepsie pourrait être due en partie à la relation bidirectionnelle entre l’épilepsie et les gènes de l’horloge biologique. Les altérations des mécanismes de l’horloge biologique augmentent la susceptibilité à l’épilepsie, tandis que les crises peuvent perturber l’horloge interne (Re et al.,2020). Un courant nettement plus élevé est nécessaire pour induire des crises d’épilepsie maximales et généralisées chez les souris de type sauvage (TS) pendant la phase obscure de leur cycle diurne, comparativement à la phase lumineuse. Ce rythme est aboli chez les souris Bmal1 KO, qui présentent également des seuils épileptogènes significativement plus bas en permanence par rapport à leurs homologues TS (Gerstner et al.,2014).
De même, l’inactivation conditionnelle de Bmal1 dans les neurones du gyrus denté a augmenté la susceptibilité aux crises induites par la pilocarpine chez la souris (Wu et al.,2021).L’expression de BMAL1 dans l’hippocampe diminue au fil du temps chez les rats traités à la pilocarpine lorsqu’ils commencent à développer des crises spontanées, ce qui suggère que BMAL1 joue également un rôle dans l’épileptogenèse (Matos et al.,2018). Il a été constaté que les niveaux de la protéine BMAL1 étaient diminués dans le gyrus denté et la région CA1 des souris atteintes d’épilepsie du lobe temporal (Wu et al.,2021). Les mutations du récepteur orphelin alpha apparenté à RAR (RORAgène, qui code pour un activateur de la transcription Bmal1l, ont été liées à un trouble du développement intellectuel avec ou sans épilepsie ou ataxie cérébelleuse (IDDECA) (Guissart et al.,2018).
La suppression du gène Horloge dans les neurones pyramidaux corticaux chez la souris entraîne des décharges épileptiformes dans les neurones excitateurs ainsi qu’une diminution du seuil épileptogène (Li et al.,2017). L’analyse par PCR quantitative en temps réel (qPCR) a révélé une perte de l’expression rythmique de CLOCK et une diminution des niveaux de son transcrit dans un modèle de rat post-état de mal épileptique (Santos et al.,2015). L’ARN Horloge et les protéines sont également sous-régulés dans les tissus cérébraux réséqués chez les patients atteints d’épilepsie du lobe temporal (Li et al.,2017).
Un autre gène d’horloge oscillante, Per1, est régulée à la hausse dans l’hippocampe à la suite de crises induites électriquement et par l’acide kaïnique chez la souris (Eun et al.,2011). Une étude a mis en évidence une altération de l’expression rythmique de PER1, PER2 et PER3 dans un modèle de rat de crises induites par la pilocarpine (Santos et al.,2015). Cependant, une étude ultérieure a révélé une augmentation de l’expression de PER1 et une diminution de l’expression de PER2 avant le développement de crises spontanées, tandis que l’expression de PER3 restait inchangée (Matos et al.,2018).
En conclusion, le sommeil et la phase circadienne ont des effets directs sur la respiration péri-ictale et potentiellement sur le développement de l’épilepsie elle-même.
Troubles du sommeil, troubles respiratoires du sommeil (TRS) et épilepsie
Privation de sommeil/troubles du sommeil
Outre les crises nocturnes, les patients épileptiques présentent également une prévalence plus élevée de troubles du sommeil que les individus sains (Vaughn et D’cruz,2004). De nombreuses études menées au cours des 30 dernières années ont montré à maintes reprises que les adultes épileptiques ont 2 à 3 fois plus de risques de souffrir de troubles du sommeil/éveil que la population générale (Grigg-Damberger et Foldvary-Schaefer, 2021). Les patients atteints d’épilepsie du lobe temporal présentent une efficacité du sommeil réduite et davantage d’éveils que ceux atteints d’épilepsie du lobe frontal (Crespel et al.,2000). De plus, la stimulation de l’amygdale diminue le sommeil paradoxal chez les animaux de laboratoire, et la privation sélective de sommeil paradoxal accélère le processus de stimulation (Cohen et Dement,1970 ; Tanaka et Naquet,1975). La souris Scn1aR1407X/+ présente des anomalies dans la régulation du sommeil circadien, notamment un rythme fragmenté du sommeil NREM et une période circadienne de sommeil allongée (Sanchez et al.,2019).
Les troubles du sommeil ou des crises nocturnes fréquentes peuvent entraîner une privation de sommeil.
La privation de sommeil elle-même peut induire des crises et des pointes interictales (Mattson et al.,1965 ; Pratt et al.,1968 ; Malow et al.,2000b ; Konduru et al.,2021). Chez les chats ayant subi une stimulation de l’amygdale, une privation aiguë de sommeil réduit le seuil de crise et de décharge post-crise (Shouse et Sterman,1982).
Cependant, une privation de sommeil plus prolongée augmente leur susceptibilité aux crises d’épilepsie induites par stimulation électrique et par la pénicilline, quel que soit l’état de sommeil (Shouse,1988).
De plus, lorsqu’on a administré à des rats sensibilisés une micro-injection d’un agoniste cholinergique dans la formation réticulaire pontique pour améliorer le sommeil paradoxal, on a observé une augmentation significative du seuil de courant nécessaire pour déclencher des décharges post-potentielles dans l’amygdale (Kumar et al.,2007). Des études sur la privation de sommeil chez des individus sains ont montré une hypertension et une augmentation de l’activité du système nerveux sympathique après des nuits où le sommeil était inférieur à 5 h (Lusardi et al.,1996 ; Tochikubo et al.,1996 ; Gangwisch et al.,2006).
Ainsi, le manque de sommeil peut non seulement aggraver les crises elles-mêmes, mais aussi rendre les patients plus vulnérables aux atteintes du système nerveux autonome induites par les crises.
TRS
Jusqu’à 9 à 11 % des patients adultes atteints d’épilepsie présentent un syndrome d’apnée du sommeil ((Vendrame et al., 2014 ; Popkirov et al., 2019).Ce chiffre atteint 40 % chez les enfants épileptiques (Kaleyias et al., 2008).Une étude de cas récente a mis en lumière le cas d’un patient de sexe masculin présentant des antécédents de crises tonico-cloniques généralisées secondaires et une respiration nocturne paroxystique. Ce patient a connu des épisodes d’arrêt respiratoire associés à un bruit expiratoire atypique, principalement pendant le sommeil paradoxal ou lors de la transition entre le sommeil paradoxal et le sommeil lent, malgré une absence de crises pendant un an (Künstler et al., 2022).
L’apnée obstructive du sommeil (AOS) est une forme relativement courante de troubles respiratoires du sommeil, dans laquelle les voies aériennes supérieures s’affaissent, empêchant la ventilation. L’apnée qui en résulte provoque une réponse d’éveil qui permet un repositionnement et la reprise des échanges gazeux (Butler et al., 2015).La prévalence exacte de l’apnée obstructive du sommeil (AOS) chez les personnes épileptiques ne fait pas encore l’objet d’un consensus.
Popkirov et al. estiment que 7 % des patients épileptiques présentent une AOS légère à modérée (Popkirov et al., 2019).Une étude polysomnographique distincte postule qu’un tiers des patients atteints d’épilepsie pharmacorésistante et candidats à une chirurgie de l’épilepsie souffrent également d’apnée obstructive du sommeil (Malow et al., 2000b). Ce résultat est également plus proche d’une estimation issue d’une méta-analyse de 2017, qui a déterminé que la prévalence de l’apnée obstructive du sommeil (AOS) légère à sévère chez les patients épileptiques était de 33,4 %, soit 2,4 fois plus élevée que chez les sujets témoins sains (Lin et al., 2017).Cette même méta-analyse a révélé que la prévalence de l’apnée obstructive du sommeil chez les patients atteints d’épilepsie réfractaire n’était pas supérieure à la prévalence globale de l’apnée obstructive du sommeil chez les patients épileptiques (Lin et al., 2017).Les patients atteints d’épilepsie généralisée présentent une apnée obstructive du sommeil plus sévère que ceux atteints d’épilepsie focale. Les deux populations ont rapporté des degrés similaires de somnolence diurne anormale.Des degrés similaires de somnolence diurne anormale ont été rapportés dans les deux populations. Un âge plus avancé, un indice de masse corporelle (IMC) plus élevé et des antécédents d’hypertension sont également associés à une apnée obstructive du sommeil (AOS) plus sévère (Scharf et al., 2020).
L’incidence de l’apnée obstructive du sommeil chez les personnes non épileptiques est plus élevée chez les hommes que chez les femmes (4 % chez les hommes, 2 % chez les femmes) (Block et al.,1979 ; Young et al., 1993). Les hommes sont également beaucoup plus susceptibles de présenter une désaturation en O2 pendant l’apnée que les femmes (Block et al.,1979). Chez les patients épileptiques, les hommes sont environ trois fois plus susceptibles de souffrir d’apnée obstructive du sommeil que les femmes (Lin et al., 2017).
La durée des apnées obstructives tend à augmenter au cours d’une nuit de sommeil (Montserrat et al.,1996 ; Butler et al., 2015). Il est suggéré que cela est dû à un atténuation de la réponse d’éveil au CO2 au cours de la nuit, entraînant des périodes d’hypercapnie plus longues avant l’éveil (Montserrat et al.,1996). Il est possible que cette augmentation de l’apnée obstructive du sommeil chez les patients épileptiques soit due à une diminution intrinsèque de la chimiosensibilité dans le cerveau épileptique. Les adolescents obèses atteints d’apnée obstructive du sommeil présentent une augmentation de la réponse chimio-sensible à l’hypoxie (HCVR) à l’état d’éveil et une diminution de cette HCVR pendant le sommeil (Yuan et al.,2012).
Il existe également des composantes circadiennes endogènes contribuant à la prolongation des événements respiratoires nocturnes. Aux phases circadiennes correspondant au début de la matinée, la durée des apnées et des hypopnées est généralement plus longue, mais l’indice d’apnée-hypopnée (IAH), qui mesure la gravité du syndrome d’apnées obstructives du sommeil (SAOS), est faible.
En revanche, en fin d’après-midi et en début de soirée, la durée des événements est courte et l’IAH est élevé. Les événements survenant pendant le sommeil paradoxal ont également tendance à être 14 % plus longs que ceux survenant après le sommeil lent (Butler et al., 2015).
La comorbidité de l’épilepsie et de l’apnée obstructive du sommeil peut augmenter l’incidence des arythmies et accroître le risque de mort subite cardiaque chez le patient (Gami et Somers, 2008 ; Gami et al., 2013). Les patients atteints d’apnée obstructive du sommeil subissent une perturbation du système nerveux autonome pendant le sommeil (Adlakha et Shepard,1998), qui peut être davantage déséquilibré par les crises. Bien qu’aucune corrélation directe entre l’apnée obstructive du sommeil (AOS) et la mort subite inattendue en épilepsie (SUDEP) n’ait été identifiée, des scores SUDEP-7 révisés plus élevés [présence de crises au cours des 12 derniers mois, en particulier de crises tonico-cloniques généralisées (CTCG), durée plus longue de l’épilepsie, nombre accru de symptômes aigus et QI plus faible/troubles cognitifs plus importants] sont associés à une SUDEP probable (Phabphal et al.,2021).
L’apnée obstructive du sommeil (AOS) diminue la durée du sommeil nocturne, ce qui peut entraîner une privation de sommeil supplémentaire. Cette privation de sommeil est particulièrement dangereuse pour les personnes épileptiques car elle peut avoir un effet épileptogène (Nobili et al., 2011 ; Popkirov et al., 2019).Il s’ensuit que lorsque les patients épileptiques atteints d’apnée obstructive du sommeil étaient traités par pression positive continue (PPC), ils présentaient un meilleur contrôle des crises que leurs pairs non traités (Lin et al., 2017).
La stimulation du nerf vague (SNV) est une technique utilisée pour traiter l’épilepsie réfractaire.via un dispositif de neuro-stimulation. Bien que ces dispositifs aient permis de réduire la fréquence et la gravité des crises, il n’existe pas de preuves concluantes indiquant que la stimulation du nerf vague (SNV) diminue le risque de mort subite inattendue en épilepsie (SUDEP) (Annegers et al.,1998 ; Ryvlin et al., 2018).Il existe cependant des preuves que l’activation du VNS pendant le sommeil peut induire une SAOS légère ou aggraver une SAOS préexistante. L’activation du VNS pendant le sommeil est également liée à une diminution du VC et de la SaO2, à une augmentation de la fréquence respiratoire et de l’IAH, ainsi qu’à une somnolence diurne excessive (Malow et al., 2000a ; Holmes et al.,2003 ; Marzec et al., 2003 ; Zambrelli et al.,2016 ; Somboon et al.,2019 ; Kim et al.,2022).
Une étude récente a également indiqué que la pente de la réponse ventriculaire gauche (RVG) est atténuée chez les patients présentant une stimulation du nerf vague active (Sainju et al., 2021).Des données suggèrent que l’exacerbation de l’apnée obstructive du sommeil après la stimulation du nerf vague est due à une réduction de l’espace glottique ou à un manque de coordination laryngo-respiratoire (Zambrelli et al., 2016).Ceci est important car les patients atteints d’épilepsie réfractaire présentent un risque accru de SUDEP (mort subite inattendue en épilepsie) et sont plus susceptibles d’opter pour la stimulation du nerf vague (SNV) comme méthode de contrôle des crises. En résumé, les personnes épileptiques sont plus susceptibles de souffrir de troubles du sommeil et de syndrome d’apnées du sommeil (SAS), ce qui peut influencer directement la fréquence des crises. De plus, un traitement courant de l’épilepsie réfractaire semble aggraver le SAS chez ces patients.
Mécanismes des neurotransmetteurs
Les mécanismes sous-jacents aux effets du sommeil et du rythme circadien sur la respiration dans l’épilepsie. Bien que ces mécanismes ne soient pas encore pleinement compris, de nombreux neurotransmetteurs et molécules de signalisation ont été impliqués. Par exemple, le neurotransmetteur monoaminergique sérotonine (5-HT) joue un rôle important dans la régulation du cycle veille-sommeil et la respiration (Jouvet, 1999 ; Richerson,2004 ; Hodges et al.,2009 ; Ptak et al.,2009 ; Hodges et Richerson, 2010 ; Depuy et al.,2011 ; Buchanan, 2013 ; Iwasaki et al., 2018 ; Smith et al.,2018). Elle est également fortement impliquée dans la physiopathologie de l’épilepsie et de la SUDEP (Bagdy et al.,2007 ; Richerson et Buchanan,2011 ; Richerson, 2013 ; Feng et Faingold, 2017 ; Li et Buchanan, 2019 ; Petrucci et al.,2020). Le tonus sérotoninergique est modulé à la fois en fonction de l’état de sommeil et de la phase circadienne, avec un nadir pendant la nuit et le sommeil (Mcginty et Harper, 1976 ; Rosenwasser et al.,1985 ; Agren et al.,1986 ; Rao et al.,1994 ; Sakai et Crochet,2001 ; Mateos et al., 2009 ; Sakai, 2011 ; Purnell et al., 2018). Il a été démontré que les neurones 5-HT du mésencéphale et du raphé médullaire sont fortement chimiosensibles (Larnicol et al.,1994 ; Richerson,1995,2004 ; Wang et al.,1998 ; Severson et al.,2003). Il est probable que les neurones 5-HT du bulbe rachidien interviennent dans l’augmentation de la respiration en réponse à une hausse du CO2, tandis que les neurones 5-HT du mésencéphale interviennent dans les réponses non respiratoires au CO2, telles que l’éveil (Richerson,2004 ; Buchanan et Richerson, 2010 ; Buchanan et al., 2015 ; Kaur et al., 2020). L’activité des neurones 5-HT du raphé médullaire est nettement réduite pendant la période ictale et post-ictale, ce qui coïncide avec une dépression respiratoire sévère (Zhan et al.,2016). De plus, des taux sériques de 5-HT post-critiques plus faibles ont été associés à l’apnée centrale post-critique (Murugesan et al., 2019).De nombreuses études ont démontré qu’un prétraitement par agents sérotoninergiques avant le déclenchement d’une crise d’épilepsie peut atténuer ce dysfonctionnement respiratoire. L’incidence de l’IRA-S chez les souris DBA/2 peut être réduite par l’administration de fluoxétine, un inhibiteur sélectif de la recapture de la 5-HT (ISRS), avant l’induction de la crise (Tupal et Faingold, 2001).2019). Une découverte similaire a été faite chez les souris DBA/1, où il a également été constaté que la fluoxétine réduisait le S-IRA sans augmenter la ventilation basale ni la réponse ventilatoire à 7 % de CO2 (Zeng et al.,2015 ; Feng et Faingold,2017). D’autres agents sérotoninergiques, comme la fenfluramine, peuvent bloquer sélectivement la S-IRA sans influencer le comportement convulsif (Feng et Faingold,2017 ; Tupal et Faingold, 2019).
Une autre molécule de signalisation monoaminergique liée à la régulation du sommeil/éveil, à la respiration et à l’épilepsie est la noradrénaline (NE ; Hobson et al., 1975 ; Aston-Jones et Bloom,1981 ; Foote et al.,1983).
Les concentrations plasmatiques de NE sont significativement plus faibles pendant le sommeil nocturne que pendant l’éveil (Linsell et al.,1985). Tout comme la 5-HT, la NE présente également un rythme circadien, les concentrations les plus faibles étant observées pendant la nuit (Morgan et al.,1973 ; Agren et al.,1986 ; Cagampang et Inouye, 1994). L’atomoxétine, un inhibiteur de la recapture de la noradrénaline (IRN), supprime l’arrêt respiratoire induit par les crises après des crises audiogènes chez les souris DBA/1 (Zhang et al.,2017 ; Zhao et al., ju2017) ainsi que les crises MES (Kruse et al.,2019). Un autre inhibiteur de la recapture de la neuraminidase (IRN), la réboxétine, et l’inhibiteur double de la recapture de la 5-HT/NE (IRSN), la duloxétine, sont également capables de supprimer l’arrêt respiratoire consécutif aux crises d’épilepsie induites par le MES (Kruse et al.,2019). Plus récemment, des données ont montré que l’activation sélective du récepteur noradrénergique α2 suffit à supprimer la S-IRA chez les souris DBA/1 (Zhang et al.,2021).
L’orexine, un neuropeptide excitateur, est également impliquée dans le sommeil et l’éveil et constitue une substance favorisant l’éveil (Sakurai,2007 ; Bonnavion et De Lecea,2010 ; Nattie et Li,2012). L’orexine présente une forte variation circadienne diurne. Ce rythme a été mesuré dans le liquide céphalo-rachidien (LCR) et l’hypothalamus de rats, avec une variation encore plus marquée dans le LCR de rats plus âgés (Yoshida et al.,2001 ; Desarnaud et al.,2004).Ce rythme circadien robuste est probablement dû en partie aux projections denses que reçoivent les neurones à orexine du noyau suprachiasmatique (NSC), le principal oscillateur circadien du cerveau (Saper et al.,2005). Les neurones à orexine contribuent également à la fonction respiratoire, notamment grâce à l’innervation orexinergique des noyaux sérotoninergiques et noradrénergiques (Kuwaki,2008 ; Inutsuka et Yamanaka,2013).On pense que l’orexine joue un rôle proconvulsivant dans l’épilepsie, bien qu’il existe une certaine divergence concernant les effets des orexines et de leurs antagonistes sur l’activité épileptique.Kcna1Chez les souris mutantes nulles, l’antagoniste double des récepteurs de l’orexine (DORA), l’almorexant, diminue l’incidence des crises graves, améliore la saturation en O2 et augmente la longévité globale (Roundtree et al.,2016 ; Iyer et al.,2020).
L’adénosine, un neuromodulateur inhibiteur, est libérée en grande quantité lors des crises d’épilepsie (pendantet Spencer,1992 ; Berman et al.,2000 ; Van Gompel et al.,2014). Il s’agit probablement d’un mécanisme d’arrêt des crises (Shen et al.,2010 ; Purnell et al.,2021a). Contrairement à la 5-HT, à la NE et à l’orexine, l’adénosine favorise le sommeil et diminue l’éveil (Feldberg et Sherwood,1954 ; Buday et al.,1961 ; Haulicǎ et al.,1973 ; Huber et al.,2004).Ainsi, les niveaux d’adénosine augmentent pendant l’éveil et diminuent pendant le sommeil (Porkka-Heiskanen et al.,2000 ; Bjorness et Greene,2009). L’adénosine a un effet inhibiteur sur la respiration, provoquant principalement une réduction de la fréquence et du volume courant (Eldridge et al.,1984 ; Lagercrantz et al.,1984 ; Wessberg et al.,1984).L’accumulation et l’élimination de l’adénosine sont régulées de manière circadienne (Cornélissen et al.,1985 ; Chagoya De Sánchez et al.,1993 ; Huston et al.,1996). L’hypothèse de l’adénosine dans la SUDEP a été proposée pour la première fois en 2010, lorsque Shen et al. ont constaté qu’une augmentation du tonus d’adénosine dans un modèle d’épilepsie à l’acide kaïnique supprimait l’activité épileptique, mais provoquait paradoxalement la mort lorsque des crises survenaient (Shen et al.,2010). Cette hypothèse postule qu’une forte libération d’adénosine survient lors d’une crise d’épilepsie, constituant un mécanisme d’arrêt. Cependant, cette augmentation importante d’adénosine extracellulaire peut entraîner une dépression respiratoire pouvant conduire à une insuffisance respiratoire terminale (Shen et al.,2010 ; Purnell et al.,2021a).
Cette liste des principales molécules de signalisation impliquées dans la fonction respiratoire et la SUDEP est loin d’être exhaustive. Toutefois, ces neuro-modulateurs présentent un intérêt particulier dans le domaine de la SUDEP, et leur rôle complexe dans la régulation du cycle veille-sommeil, la respiration et les crises d’épilepsie en fait d’excellents candidats pour une intervention thérapeutique.
Conclusions
La mort subite inattendue en épilepsie (SUDEP) est un phénomène complexe et dévastateur, dont les mécanismes sous-jacents commencent à peine à être élucidés. Le moment de la journée et l’état de sommeil pendant lesquels surviennent les crises sont incontestablement des facteurs de risque supplémentaires pour les patients épileptiques. Bien que nuit et sommeil soient souvent liés, il est crucial de reconnaître qu’ils ne sont pas synonymes et qu’ils présentent chacun leurs propres facteurs de risque, issus de mécanismes communs et distincts. L’insuffisance respiratoire est un facteur déclenchant majeur de la mort subite induite par une crise. Les neurones monoaminergiques, notamment la sérotonine (5-HT), la noradrénaline (NE) et l’orexine, jouent un rôle crucial dans la fonction respiratoire et ont des propriétés protectrices contre les crises. Leur concentration diminue pendant la nuit et encore davantage pendant le sommeil. Ceci pourrait expliquer pourquoi les crises survenant au réveil ont tendance à être plus longues et à provoquer une insuffisance respiratoire plus sévère. D’autres molécules de signalisation, comme l’adénosine, pourraient jouer un rôle encore plus complexe dans la physiopathologie de la SUDEP, contribuant à l’insuffisance respiratoire lors de l’arrêt des crises. De nombreuses victimes de SUDEP sont retrouvées en position ventrale dans leur lit, ce qui suggère que la détresse respiratoire a été amplifiée par une obstruction des voies aériennes. Comme les patients sont plus souvent seuls la nuit, les chances de succès d’une intervention sont faibles.
Ainsi, bien que le sommeil et la nuit semblent présenter un risque propre de SUDEP, leur concomitance contribue fortement à la conjonction de facteurs qui conduit finalement à la mort subite induite par une crise d’épilepsie. Néanmoins, nous espérons que cette analyse permettra de comprendre que l’état de sommeil et le moment de la journée sont des facteurs à considérer indépendamment lors de l’élaboration de stratégies préventives visant à atténuer la gravité des troubles respiratoires provoqués par les crises d’épilepsie.
Références
Adlakha, A., and Shepard, J. W. (1998). Cardiac arrhythmias during normal sleep and in obstructive sleep apnea syndrome. Sleep Med. Rev. 2, 45–60. doi : 10.1016/s1087-0792(98)90053-3
Adolph, E. F. (1969). Regulations during survival without oxygen in infant mammals. Respir. Physiol. 7, 356–368. doi : 10.1016/0034-5687(69) 90019-x
Agren, H., Koulu, M., Saavedra, J. M., Potter, W. Z., and Linnoila, M. (1986).
Circadian covariation of norepinephrine and serotonin in the locus coeruleus and dorsal raphe nucleus in the rat. Brain Res. 397, 353–358. doi : 10.1016/0006- 8993(86)90638-4
Aiba, I., and Noebels, J. L. (2015). Spreading depolarization in the brainstem mediates sudden cardiorespiratory arrest in mouse SUDEP models. Sci. Transl.
Med. 7:282ra46. doi : 10.1126/scitranslmed.aaa4050
Ainslie, P. N., Lucas, S. J. E., and Burgess, K. R. (2013). Breathing and sleep at high altitude. Respir. Physiol. Neurobiol. 188, 233–256. doi : 10.1016/j.resp.2013. 05.020
Ali, A., Wu, S., Issa, N. P., Rose, S., Towle, V. L., Warnke, P., et al. (2017).
Association of sleep with sudden unexpected death in epilepsy. Epilepsy Behav. 76, 1–6. d
oi : 10.1016/j.yebeh.2017.08.021
Annegers, J. F., Coan, S. P., Hauser, W. A., Leestma, J., Duffell, W., and Tarver, B. (1998). Epilepsy, vagal nerve stimulation by the ncp system, mortality and sudden, unexpected, unexplained death. Epilepsia 39, 206–212. doi : 10.1111/j.1528-1157. 1998.tb01360.x
Aserinsky, E., and Kleitman, N. (1953). Regularly occurring periods of eye motility and concomitant phenomena, during sleep. Science 118, 273–274. doi : 10.1126/science.118.3062.273
Aston-Jones, G., and Bloom, F. E. (1981). Activity of norepinephrinecontaining locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. J. Neurosci. 1, 876–886. doi : 10.1523/JNEUROSCI.01-08-00 876.1981
Bagdy, G., Kecskemeti, V., Riba, P., and Jakus, R. (2007). Serotonin and epilepsy. J. Neurochem. 100, 857–873. doi : 10.1111/j.1471-4159.2006. 04277.x
Bando, H., Nishio, T., Van Der Horst, G. T. J., Masubuchi, S., Hisa, Y., and Okamura, H. (2007). Vagal regulation of respiratory clocks in mice. J. Neurosci. 27, 4359–4365. doi : 10.1523/JNEUROSCI.4131-06.2007
Bateman, L. M., Li, C. S., and Seyal, M. (2008). Ictal hypoxemia in localizationrelated epilepsy : analysis of incidence, severity and risk factors. Brain 131, 3239–3245. doi : 10.1093/brain/awn277
Bayne, L. L., and Simon, R. P. (1981). Systemic and pulmonary vascular pressures during generalized seizures in sheep. Ann. Neurol. 10, 566–569. doi : 10.1002/ana. 410100613
Bazil, C. W., and Walczak, T. S. (1997). Effects of sleep and sleep stage on epileptic and nonepileptic seizures. Epilepsia 38, 56–62. doi : 10.1111/j.1528-1157. 1997.tb01077.x
Berman, R. F., Fredholm, B. B., Aden, U., and O’connor, W. T. (2000). Evidence for increased dorsal hippocampal adenosine release and metabolism during pharmacologically induced seizures in rats. Brain Res. 872, 44–53. doi : 10.1016/s0006-8993(00)02441-0
Berssenbrugge, A., Dempsey, J., Iber, C., Skatrud, J., and Wilson, P. (1983). Mechanisms of hypoxia-induced periodic breathing during sleep in humans. J. Physiol. 343, 507–524. doi : 10.1113/jphysiol.1983.
sp014906
Berthon-Jones, M., and Sullivan, C. E. (1982). Ventilatory and arousal responses to hypoxia in sleeping humans. Am. Rev. Respir. Dis. 125, 632–639. doi : 10.1164/arrd.1982.125.6.632
Berthon-Jones, M., and Sullivan, C. E. (1984). Ventilation and arousal responses to hypercapnia in normal sleeping humans. J. Appl. Physiol. Respir. Environ. Exerc.Physiol. 57, 59–67. doi : 10.1152/jappl.1984.57.1.59
Birchfield, R. I., Sieker, H. O., and Heyman, A. (1959). Alterations in respiratory function during natural sleep. J. Lab. Clin. Med. 54, 216–222.
Bjorness, T. E., and Greene, R. W. (2009). Adenosine and sleep. Curr.Neuropharmacol. 7, 238–245. doi : 10.2174/157015909789152182
Block, A. J., Boysen, P. G., Wynne, J. W., and Hunt, L. A. (1979). Sleep apnea,hypopnea and oxygen desaturation in normal subjects. N. Engl. J. Med. 300,513–517. doi : 10.1056/NEJM197903083001001
Blum, A. S. (2009). Respiratory physiology of seizures. J. Clin. Neurophysiol. 26,309–315.
doi : 10.1097/WNP.0b013e3181b7f14d
Bonnavion, P., and De Lecea, L. (2010). Hypocretins in the control of sleep and wakefulness. Curr. Neurol. Neurosci. Rep. 10, 174–179. doi : 10.1007/s11910-010-0101-y
Bozorgi, A., and Lhatoo, S. D. (2013). Seizures, cerebral shutdown and SUDEP.Epilepsy Curr. 13, 236–240. doi : 10.5698/1535-7597-13.5.236
Browne, K. F., Prystowsky, E., Heger, J. J., Chilson, D. A., and Zipes, D. P. (1983).Prolongation of the q-t interval in man during sleep. Am. J. Cardiol. 52, 55–59. doi : 10.1016/0002-9149(83)90068-1
Bryan, A. C., and Muller, N. L. (1980). Lung mechanics and gas exchange duringsleep. Sleep 3, 401–416. doi : 10.1093/sleep/3.3-4.401
Buchanan, G. F. (2013). Timing, sleep and respiration in health and disease. Prog.Mol. Biol. Transl. Sci. 119, 191–219. doi : 10.1016/B978-0-12-396971-2.00008-7
Buchanan, G. F. (2019). Impaired CO2-induced arousal in SIDS and SUDEP.Trends Neurosci. 42, 242–250. doi : 10.1016/j.tins.2019.02.002
Buchanan, G. F., Murray, N. M., Hajek, M. A., and Richerson, G. B.(2014). Serotonin neurones have anti-convulsant effects and reduce seizureinduced mortality. J. Physiol. 592, 4395–4410.
doi : 10.1113/jphysiol.2014.277574
Buchanan, G. F., and Richerson, G. B. (2010). Central serotonin neurons are required for arousal to CO2. Proc. Natl. Acad. Sci. U S A 107, 16354–16359. doi : 10.1073/pnas.1004587107
Buchanan, G. F., Smith, H. R., Macaskill, A., and Richerson, G. B. (2015). 5-HT2A receptor activation is necessary for CO2-induced arousal. J. Neurophysiol.114, 233–243. doi : 10.1152/jn.00213.2015
Buday, P. V., Carr, C. J., and Miya, T. S. (1961). A pharmacologic study of some nucleosides and nucleotides. J. Pharm Pharmacol. 13, 290–299. doi : 10.1111/j. 2042-7158.1961.tb11826.x
Budde, R. B., Arafat, M. A., Pederson, D. J., Lovick, T. A., Jefferys, J. G. R., and Irazoqui, P. P. (2018). Acid reflux induced laryngospasm as a potential mechanism of sudden death in epilepsy. Epilepsy Res. 148, 23–31. doi : 10.1016/j.eplepsyres.2018.10.003
Bulow, K. (1963). Respiration and wakefulness in man. Acta Physiol. Scand.Suppl. 209, 1–110.
Butler, M. P., Smales, C., Wu, H., Hussain, M. V., Mohamed, Y. A., Morimoto, M.,et al. (2015). The circadian system contributes to apnea lengthening across the night in obstructive sleep apnea. Sleep 38, 1793–1801. doi : 10.5665/sleep.5166
Cagampang, F. R. A., and Inouye, S.-I. T. (1994). Diurnal and circadian changes of serotonin in the suprachiasmatic nuclei : regulation by light and an endogenous pacemaker. Brain Res. 639, 175–179. doi : 10.1016/0006-8993(94) 91780-9
Chagoya De Sánchez, V., Múñoz, R. H., Suárez, J., Vidrio, S., Yáñez, L., and Múñoz, M. D. (1993). Day-night variations of adenosine and its metabolizing enzymes in the brain cortex of the rat — possible physiological significance for the energetic homeostasis and the sleep-wake cycle. Brain Res. 612, 115–121.
doi : 10.1016/0006-8993(93)91651-8
Chen, Z., Brodie, M. J., Liew, D., and Kwan, P. (2018). Treatment outcomes in patients with newly diagnosed epilepsy treated with established and new antiepileptic drugs : a 30-year longitudinal cohort study. JAMA Neurol. 75, 279–286. doi : 10.1001/jamaneurol.2017.3949
Cherniack, N. S. (1981). Respiratory dysrhythmias during sleep. N. Engl. J. Med. 305, 325–330. doi : 10.1056/NEJM198108063050606
Cherniack, N. S. (1999). Apnea and periodic breathing during sleep. N. Engl. J. Med. 341, 985–987. doi : 10.1056/NEJM199909233411310
Chrobok, L., Northeast, R. C., Myung, J., Cunningham, P. S., Petit, C., and Piggins, H. D. (2020). Timekeeping in the hindbrain : a multi-oscillatory circadian centre in the mouse dorsal vagal complex. Commun. Biol. 3:225.
doi : 10.1038/s42003-020-0960-y
Cohen, H. B., and Dement, W. C. (1970). Prolonged tonic convulsions in REM deprived mice. Brain Res. 22, 421–422. doi : 10.1016/0006-8993(70)90487-7
Cornélissen, G., Touitou, Y., Tritsch, G., Bogdan, A., Auzéby, A., Reinberg, A., et al. (1985). Circadian rhythms of adenosine deaminase activity in human erythrocytes : a transverse study on young, elderly and senile demented subjects. La Ricerca in Clin. Lab. 15, 365–374. doi : 10.1007/BF03029152
Crespel, A., Coubes, P., and Baldy-Moulinier, M. (2000). Sleep influence on seizures and epilepsy effects on sleep in partial frontal and temporal lobe epilepsies. Clin. Neurophysiol. 111, S54–S59. doi : 10.1016/s1388-2457(00) 00402-8
Del Negro, C. A., Funk, G. D., and Feldman, J. L. (2018). Breathing matters. Nat. Rev. Neurosci. 19, 351–367. doi : 10.1038/s41583-018-0003-6
Depuy, S. D., Kanbar, R., Coates, M. B., Stornetta, R. L., and Guyenet, P. G. (2011). Control of breathing by raphe obscurus serotonergic neurons in mice. J. Neurosci. 31, 1981–1990. doi : 10.1523/JNEUROSCI.4639
– 10.2011
Desarnaud, F., Murillo-Rodriguez, E., Lin, L., Xu, M., Gerashchenko, D., Shiromani, S. N., et al. (2004). The diurnal rhythm of hypocretin in young and old f344 rats. Sleep 27, 851–856. doi : 10.1093/sleep/27.5.851
Devinsky, O., Hesdorffer, D. C., Thurman, D. J., Lhatoo, S., and Richerson, G. (2016). Sudden unexpected death in epilepsy : epidemiology, mechanisms and prevention. Lancet Neurol. 15, 1075–1088. doi : 10.1016/S1474-4422(16) 30158-2
Dhaibar, H., Gautier, N. M., Chernyshev, O. Y., Dominic, P., and Glasscock, E. (2019). Cardiorespiratory profiling reveals primary breathing dysfunction in kcna1-null mice : implications for sudden unexpected death in epilepsy. Neurobiol. Dis. 127, 502–511. doi : 10.1016/j.nbd.2019.04.006
Dlouhy, B. J., Gehlbach, B. K., Kreple, C. J., Kawasaki, H., Oya, H., Buzza, C., et al. (2015). Breathing inhibited when seizures spread to the amygdala and upon amygdala stimulation. J. Neurosci. 35, 10281–10289. doi : 10.1523/JNEUROSCI. 0888-15.2015
Douglas, N. J. (2000). Sleep and breathing. Schweiz. Med. Wochenschr. 130, 272–276.
Douglas, N. J., White, D. P., Pickett, C. K., Weil, J. V., and Zwillich, C. W. (1982a). Respiration during sleep in normal man. Thorax 37, 840–844. doi : 10.1136/thx.37. 11.840
Douglas, N. J., White, D. P., Weil, J. V., Pickett, C. K., Martin, R. J., Hudgel, D. W., et al. (1982b). Hypoxic ventilatory response decreases during sleep in normal men. Am. Rev. Respir. Dis. 125, 286–289. doi : 10.1164/arrd.1982.125.3.286
Douglas, N. J., White, D. P., Weil, J. V., Pickett, C. K., and Zwillich, C. W. (1982c). Hypercapnic ventilatory response in sleeping adults. Am. Rev. Respir. Dis. 126, 758–762. doi : 10.1164/arrd.1982.126.5.758
During, M. J., and Spencer, D. D. (1992). Adenosine : a potential mediator of seizure arrest and postictal refractoriness. Ann. Neurol. 32, 618–624. doi : 10.1002/ana.410320504
Eldridge, F. L., Millhorn, D. E., and Kiley, J. P. (1984). Respiratory effects of a long-acting analog of adenosine. Brain Res. 301, 273–280. doi : 10.1016/0006- 8993(84)91096-5
Eun, B., Kim, H. J., Kim, S. Y., Kim, T. W., Hong, S. T., Choi, K., et al. (2011). Induction of per1 expression following an experimentally induced epilepsy in the mouse hippocampus. Neurosci. Lett. 498, 110–113. doi : 10.1016/j.neulet.2011.03. 039
Feldberg, W., and Sherwood, S. L. (1954). Injections of drugs into the lateral ventricle of the cat. J. Physiol. 123, 148–167. doi : 10.1113/jphysiol.1954.sp005040
Feng, H. J., and Faingold, C. L. (2017). Abnormalities of serotonergic neurotransmission in animal models of SUDEP. Epilepsy Behav. 71, 174–180. doi : 10.1016/j.yebeh.2015.06.008
Fenik, V. B., Davies, R. O., and Kubin, L. (2005). REM sleep-like atonia of hypoglossal (XII) motoneurons is caused by loss of noradrenergic and serotonergic inputs. Am. J. Respir. Crit. Care Med. 172, 1322–1330. doi : 10.1164/rccm.200412- 1750OC
Foote, S. L., Bloom, F. E., and Aston-Jones, G. (1983). Nucleus locus ceruleus : new evidence of anatomical and physiological specificity. Physiol. Rev. 63, 844–914. doi : 10.1152/physrev.1983.63.3.844
Gami, A. S., Olson, E. J., Shen, W. K., Wright, R. S., Ballman, K. V., Hodge, D. O., et al. (2013). Obstructive sleep apnea and the risk of sudden cardiac death : a longitudinal study of 10,701 adults. J. Am. Coll. Cardiol. 62, 610–616. doi : 10.1016/j. jacc.2013.04.080
Gami, A. S., and Somers, V. K. (2008). Implications of obstructive sleep apnea for atrial fibrillation and sudden cardiac death. J. Cardiovasc. Electrophysiol. 19, 997–1003. doi : 10.1111/j.1540-8167.2008.01136.x
Gangwisch, J. E., Heymsfield, S. B., Boden-Albala, B., Buijs, R. M., Kreier, F.,
Pickering, T. G., et al. (2006). Short sleep duration as a risk factor for hypertension :
analyses of the first national health and nutrition examination survey. Hypertension
47, 833–839. doi : 10.1161/01.HYP.0000217362.34748.e0
Gerstner, J. R., Smith, G. G., Lenz, O., Perron, I. J., Buono, R. J., and Ferraro, T. N.
(2014). Bmal1 controls the diurnal rhythm and set point for electrical seizure
threshold in mice. Front. Syst. Neurosci. 8:121. doi : 10.3389/fnsys.2014.00121
Gharib, A., Sayyahi, Z., Komaki, A., Barkley, V., Sarihi, A., and MirnajafiZadeh, J. (2018). The role of 5-HT1A receptors of hippocampal CA1 region in
anticonvulsant effects of low-frequency stimulation in amygdala kindled rats.
Physiol. Behav. 196, 119–125. doi : 10.1016/j.physbeh.2018.08.025
Grigg-Damberger, M., and Foldvary-Schaefer, N. (2021). Bidirectional relationships of sleep and epilepsy in adults with epilepsy. Epilepsy Behav.116:107735. doi : 10.1016/j.yebeh.2020.107735
Guissart, C., Latypova, X., Rollier, P., Khan, T. N., Stamberger, H., Mcwalter, K.,et al. (2018). Dual molecular effects of dominant RORA mutations cause two variants of syndromic intellectual disability with either autism or cerebellar ataxia. Am. J. Hum Genet. 102, 744–759. doi : 10.1016/j.ajhg.2018.02.021
Guntheroth, W. G., and Kawabori, I. (1975). Hypoxic apnea and gasping. J. Clin.Invest. 56, 1371–1377. doi : 10.1172/JCI108217
Hajek, M. A., and Buchanan, G. F. (2016). Influence of vigilance state on physiological consequences of seizures and seizure-induced death in mice. J. Neurophysiol. 115, 2286–2293. doi : 10.1152/jn.00011.2016
Harden, C., Tomson, T., Gloss, D., Buchhalter, J., Cross, J. H., Donner, E., et al. (2017). Practice guideline summary : sudden unexpected death in epilepsy incidence rates and risk factors : report of the guideline development, dissemination and implementation subcommittee of the american academy of neurology and the american epilepsy society. Epilepsy Curr. 17, 180–187. doi : 10.5698/1535-7511.17.3.180
Haulica, I., Ababei, L., Br ˇ ani¸slteanu, D., and Topoliceanu, F. (1973). Preliminary data on the possible hypnogenic role of adenosine1. J. Neurochem. 21, 1019–1020.doi : 10.1111/j.1471-4159.1973.tb07549.x
Haxhiu, M. A., Mack, S. O., Wilson, C. G., Feng, P., and Strohl, K. P. (2003). Sleep networks and the anatomic and physiologic connections with respiratory control.Front. Biosci. 8, d946–d962. doi : 10.2741/1099
Haxhiu, M. A., Van Lunteren, E., Mitra, J., and Cherniack, N. S. (1987).Comparison of the response of diaphragm and upper airway dilating muscle activity in sleeping cats. Respir. Physiol. 70, 183–193. doi : 10.1016/0034-5687(87)90049-1
Hesdorffer, D. C., Tomson, T., Benn, E., Sander, J. W., Nilsson, L., Langan, Y., et al. (2011). Combined analysis of risk factors for SUDEP. Epilepsia 52, 1150–1159. doi : 10.1111/j.1528-1167.2010.02952.x
Hicks, J. W., and Riedesel, M. L. (1983). Diurnal ventilatory patterns in the garter snake, Thamnophis elegans. J. Comp. Physiol. 149, 503–510.doi : 10.1007/BF00690009
Hobson, J. A., Mccarley, R. W., and Wyzinski, P. W. (1975). Sleep cycle oscillation : reciprocal discharge by two brainstem neuronal groups. Science 4,55–58. doi : 10.1126/science.1094539
Hodges, M. R., and Richerson, G. B. (2010). Medullary serotonin neurons and their roles in central respiratory chemoreception. Respir. Physiol. Neurobiol. 173,256–263. doi : 10.1016/j.resp.2010.03.006
Hodges, M. R., Wehner, M., Aungst, J., Smith, J. C., and Richerson, G. B. (2009). Transgenic mice lacking serotonin neurons have severe apnea and high mortality during development. J. Neurosci. 29, 10341–10349. doi : 10.1523/JNEUROSCI. 1963-09.2009
Holmes, M. D., Chang, M., and Kapur, V. (2003). Sleep apnea and excessive daytime somnolence induced by vagal nerve stimulation. Neurology 61, 1126–1129. doi : 10.1212/01.wnl.0000086812.62554.06
Huber, R., Felice Ghilardi, M., Massimini, M., and Tononi, G. (2004). Local sleep and learning. Nature 430, 78–81. doi : 10.1038/nature02663 Huston, J. P., Haas, H. L., Boix, F., Pfister, M., Decking, U., Schrader, J., et al.
(1996). Extracellular adenosine levels in neostriatum and hippocampus during rest and activity periods of rats. Neuroscience 73, 99–107. doi : 10.1016/0306-4522(96)00021-8
Iizuka, H., Sasaki, K., Odagiri, N., Obo, M., Imaizumi, M., and Atai, H. (2010). Measurement of respiratory function using whole-body plethysmography in unanesthetized and unrestrained nonhuman primates. J. Toxicol. Sci. 35, 863–870. doi : 10.2131/jts.35.863
Inutsuka, A., and Yamanaka, A. (2013). The physiological role of orexin/hypocretin neurons in the regulation of sleep/wakefulness and neuroendocrine functions. Front. Endocrinol. (Lausanne) 4:18. doi : 10.3389/fendo.
2013.00018
Irizarry, R., Sukato, D., Kollmar, R., Schild, S., Silverman, J., Sundaram, K., et al. (2020). Seizures induce obstructive apnea in DBA/2J audiogenic seizureprone mice : lifesaving impact of tracheal implants. Epilepsia 61, e13–e16. doi : 10.1111/epi.16431
Iwasaki, K., Komiya, H., Kakizaki, M., Miyoshi, C., Abe, M., Sakimura, K., et al. (2018). Ablation of central serotonergic neurons decreased rem sleep and attenuated arousal response. Front. Neurosci. 12:535. doi : 10.3389/fnins.2018. 00535
Iyer, S. H., Aggarwal, A., Warren, T. J., Hallgren, J., Abel, P. W., Simeone, T. A., et al. (2020). Progressive cardiorespiratory dysfunction in kv1.1 knockout mice may provide temporal biomarkers of pending sudden unexpected death in epilepsy (SUDEP) : the contribution of orexin. Epilepsia 61, 572–588. doi : 10.1111/epi.16434
Jansen, N. A., Schenke, M., Voskuyl, R. A., Thijs, R. D., Van Den Maagdenberg, A., and Tolner, E. A. (2019). Apnea associated with brainstem seizures in Cacna1a (s218l) mice is caused by medullary spreading depolarization. J. Neurosci. 39, 9633–9644. doi : 10.1523/JNEUROSCI.1713-19.2019
Javaheri, S., and Guerra, L. F. (1990). Effects of domperidone and medroxyprogesterone acetate on ventilation in man. Respir. Physiol. 81, 359–370. doi : 10.1016/0034-5687(90)90116-g
Jefferys, J. G. R., Arafat, M. A., Irazoqui, P. P., and Lovick, T. A. (2019). Brainstem activity, apnea and death during seizures induced by intrahippocampal kainic acid in anaesthetized rats. Epilepsia 60, 2346–2358. doi : 10.1111/epi.16374
Jin, Z. L., Chen, X. F., Ran, Y. H., Li, X. R., Xiong, J., Zheng, Y. Y., et al. (2017). Mouse strain differences in SSRI sensitivity correlate with serotonin transporter binding and function. Sci. Rep. 7:8631. doi : 10.1038/s41598-017- 08953-4
Johnston, S. C., Siedenberg, R., Min, J. K., Jerome, E. H., and Laxer, K. D. (1997). Central apnea and acute cardiac ischemia in a sheep model of epileptic sudden death. Ann. Neurol. 42, 588–594. doi : 10.1002/ana.410420409 Jouvet, M. (1999). Sleep and serotonin an unfinished story.
Neuropsychopharmacology 21, 24S–27S. doi : 10.1016/S0893-133X(99) 00009-3
Junkermann, H., Mangold, H., Vecsei, P., and Runnebaum, B. (1982). Circadian rhythm of serum progesterone levels in human pregnancy and its relation to the rhythm of cortisol. Acta Endocrinol. (Copenh) 101, 98–104. doi : 10.1530/acta.0. 1010098
Kaleyias, J., Cruz, M., Goraya, J. S., Valencia, I., Khurana, D. S., Legido, A., et al. (2008). Spectrum of polysomnographic abnormalities in children with epilepsy. Pediatr. Neurol. 39, 170–176. doi : 10.1016/j.pediatrneurol.2008.06.002
Kaneko, K., Yamada, T., Tsukita, S., Takahashi, K., Ishigaki, Y., Oka, Y., et al. (2009). Obesity alters circadian expressions of molecular clock genes in the brainstem. Brain Res. 1263, 58–68. doi : 10.1016/j.brainres.2008.12.071
Kaur, S., De Luca, R., Khanday, M. A., Bandaru, S. S., Thomas, R. C., Broadhurst, R. Y., et al. (2020). Role of serotonergic dorsal raphe neurons in hypercapnia-induced arousals. Nat. Commun. 11:2769. doi : 10.1038/s41467-020-16518-9
Kelly, M. N., Smith, D. N., Sunshine, M. D., Ross, A., Zhang, X., Gumz, M. L.,et al. (2020). Circadian clock genes and respiratory neuroplasticity genes oscillate in the phrenic motor system. Am. J. Physiol. Regul. Integr. Comp. Physiol. 318, R1058–R1067. doi : 10.1152/ajpregu.00010.2020
Kemp, J. S., Nelson, V. E., and Thach, B. T. (1994). Physical properties of bedding that may increase risk of sudden infant death syndrome in prone-sleeping infants. Pediatr. Res. 36, 7–11. doi : 10.1203/00006450-199407001-00002
Kennedy, J. D., Hardin, K. A., Parikh, P., Li, C., and Seyal, M. (2015). Pulmonary edema following generalized tonic clonic seizures is directly associated with seizure duration. Seizure 27, 19–24.
doi : 10.1016/j.seizure.2015.02.023
Kim, Y., Bravo, E., Thirnbeck, C. K., Smith-Mellecker, L. A., Kim, S. H., Gehlbach, B. K., et al. (2018). Severe peri-ictal respiratory dysfunction is common
in dravet syndrome. J. Clin. Invest. 128, 1141–1153. doi : 10.1172/JCI94999
Kim, J. S., Lee, D. E., Bae, H., Song, J. Y., Yang, K. I., and Hong, S. B.
(2022). Effects of vagus nerve stimulation on sleep-disordered breathing, daytime
sleepiness and sleep quality in patients with drug-resistant epilepsy. J. Clin. Neurol.
18, 315–322. doi : 10.3988/jcn.2022.18.3.315
Konduru, S. S., Pan, Y.-Z., Wallace, E., Pfammatter, J. A., Jones, M. V., and
Maganti, R. K. (2021). Sleep deprivation exacerbates seizures and diminishes
GABAergic tonic inhibition. Ann. Neurol. 90, 840–844. doi : 10.1002/ana.26208
Kotsopoulos, I. A., Van Merode, T., Kessels, F. G., De Krom, M. C., and
Knottnerus, J. A. (2002). Systematic review and meta-analysis of incidence studies
of epilepsy and unprovoked seizures. Epilepsia 43, 1402–1409. doi : 10.1046/j.1528-
1157.2002.t01-1-26901.x
Krieger, J. (2005). Respiratory physiology : breathing in normal subjects. Princ.
Pract. Sleep Med. 232, 232–244. doi : 10.1016/B0-72-160797-7/50026-4
Kruse, S. W., Dayton, K. G., Purnell, B. S., Rosner, J. I., and Buchanan, G. F.
(2019). Effect of monoamine reuptake inhibition and alpha1 blockade on
respiratory arrest and death following electroshock-induced seizures in mice.
Epilepsia 60, 495–507. doi : 10.1111/epi.14652
Kubin, L. (2016). Neural control of the upper airway : respiratory and statedependent mechanisms. Compr. Physiol. 6, 1801–1850. doi : 10.1002/cphy.c160002
Kumar, G., Couper, A., O’brien, T. J., Salzberg, M. R., Jones, N. C., Rees, S. M., et al. (2007). The acceleration of amygdala kindling epileptogenesis by chronic low-dose corticosterone involves both mineralocorticoid and glucocorticoid receptors. Psychoneuroendocrinology 32, 834–842. doi : 10.1016/j.psyneuen.2007.
05.011
Künstler, E. C. S., Schwab, M., and Rupprecht, S. (2022). A 37-year-old man with structural focal epilepsy and paroxysmal nocturnal breathing arrests. Chest 161, e309–e312. doi : 10.1016/j.chest.2021.12.631
Kuo, F. S., Cleary, C. M., Loturco, J. L., Chen, X., and Mulkey, D. K. (2019). Disordered breathing in a mouse model of dravet syndrome. eLife 8:e43387. doi : 10.7554/eLife.43387
Kuo, J., Zhao, W., Li, C. S., Kennedy, J. D., and Seyal, M. (2016). Postictal immobility and generalized EEG suppression are associated with the severity of respiratory dysfunction. Epilepsia 57, 412–417. doi : 10.1111/epi.13312
Kuwaki, T. (2008). Orexinergic modulation of breathing across vigilance states. Respir. Physiol. Neurobiol. 164, 204–212. doi : 10.1016/j.resp.2008.03.011
Kwan, P., and Brodie, M. J. (2000). Early identification of refractory epilepsy. N. Engl. J. Med. 342, 314–319. doi : 10.1056/NEJM20000203 3420503
Lacuey, N., Zonjy, B., Hampson, J. P., Rani, M. R. S., Zaremba, A., Sainju, R. K., et al. (2018). The incidence and significance of periictal apnea in epileptic seizures. Epilepsia 59, 573–582. doi : 10.1111/epi.14006
Lacuey, N., Zonjy, B., Londono, L., and Lhatoo, S. D. (2017). Amygdala and hippocampus are symptomatogenic zones for central apneic seizures. Neurology 88, 701–705. doi : 10.1212/WNL.0000000000003613
Lagercrantz, H., Yamamoto, Y., Fredholm, B. B., Prabhakar, N. R., and Von Euler, C. (1984). Adenosine analogues depress ventilation in rabbit neonates. Theophylline stimulation of respiration via adenosine receptors ? Pediatr. Res. 18,
387–390. doi : 10.1203/00006450-198404000-00018
Lamberts, R. J., Thijs, R. D., Laffan, A., Langan, Y., and Sander, J. W. (2012). Sudden unexpected death in epilepsy : people with nocturnal seizures may be at highest risk. Epilepsia 53, 253–257. doi : 10.1111/j.1528-1167.2011.03360.x
Langan, Y., Nashef, L., and Sander, J. W. (2005). Case-control study of SUDEP. Neurology 64, 1131–1133. doi : 10.1212/01.WNL.0000156352. 61328.CB
Larnicol, N., Wallois, F., Berquin, P., Gross, F., and Rose, D. (1994). C-fos-like immunoreactivity in the cat’s neuraxis following moderate hypoxia or hypercapnia. J. Physiol. Paris 88, 81–88.
doi : 10.3389/fped.2022.884120
Latreille, V., Abdennadher, M., Dworetzky, B. A., Ramel, J., White, D., Katz, E., et al. (2017). Nocturnal seizures are associated with more severe hypoxemia and increased risk of postictal generalized EEG suppression. Epilepsia 58, e127–e131. doi : 10.1111/epi.13841
Leguia, M. G., Andrzejak, R. G., Rummel, C., Fan, J. M., Mirro, E. A., Tcheng, T. K., et al. (2021). Seizure cycles in focal epilepsy. JAMA Neurol. 78, 454–463. doi : 10.1001/jamaneurol.2020.5370
Li, R., and Buchanan, G. F. (2019). Scurrying to understand sudden expected death in epilepsy : insights from animal models. Epilepsy Curr. 19, 390–396. doi : 10.1177/1535759719874787
Li, Q., Li, Y., Wang, X., Qi, J., Jin, X., Tong, H., et al. (2017). Fbxl4 serves as a clock output molecule that regulates sleep through promotion of rhythmic degradation of the GABAA receptor. Curr. Biol. 27, 3616–3625.e5. doi : 10.1016/j.cub.2017.10.052
Lin, Z., Si, Q., and Xiaoyi, Z. (2017). Obstructive sleep apnoea in patients with epilepsy : a meta-analysis. Sleep Breath. 21, 263–270. doi : 10.1007/s11325-016 -1391-3
Linsell, C. R., Lightman, S. L., Mullen, P. E., Brown, M. J., and Causon, R. C. (1985). Circadian rhythms of epinephrine and norepinephrine in man. J. Clin. Endocrinol. Metab. 60, 1210–1215. doi : 10.1210/jcem-60-6-1210
Lusardi, P., Mugellini, A., Preti, P., Zoppi, A., Derosa, G., and Fogari, R. (1996). Effects of a restricted sleep regimen on ambulatory blood pressure monitoring in normotensive subjects. Am. J. Hypertens. 9, 503–505. doi : 10.1016/0895- 7061(95)00389-4
Malik, V., Smith, D., and Lee-Chiong, T., Jr. (2012). Respiratory physiology during sleep. Sleep Med. Clin. 7, 497–505. doi : 10.1016/j.jsmc.2012.06.011 Malow, B. A., Edwards, J., Marzec, M., Sagher, O., and Fromes, G. (2000a).
Effects of vagus nerve stimulation on respiration during sleep : a pilot study. Neurology 55, 1450–1454. doi : 10.1212/wnl.55.10.1450 Malow, B. A., Levy, K., Maturen, K., and Bowes, R. (2000b). Obstructive
sleep apnea is common in medically refractory epilepsy patients. Neurology 55, 1002–1007. doi : 10.1212/wnl.55.7.1002
Malow, B. A., Lin, X., Kushwaha, R., and Aldrich, M. S. (1998). Interictal spiking increases with sleep depth in temporal lobe epilepsy. Epilepsia 39, 1309–1316. doi : 10.1111/j.1528-1157.1998.tb01329.x
Marincovich, A., Bravo, E., Dlouhy, B., and Richerson, G. B. (2021). Amygdala lesions reduce seizure-induced respiratory arrest in DBA/1 mice. Epilepsy Behav. 121:106440. doi : 10.1016/j.yebeh.2019.07.041
Marzec, M., Edwards, J., Sagher, O., Fromes, G., and Malow, B. A. (2003). Effects of vagus nerve stimulation on sleep-related breathing in epilepsy patients. Epilepsia 44, 930–935. doi : 10.1046/j.1528-1157.2003.56202.x
Matos, H. D. C., Koike, B. D. V., Pereira, W. D. S., De Andrade, T. G., Castro, O. W., Duzzioni, M., et al. (2018). Rhythms of core clock genes and spontaneous locomotor activity in post-status epilepticus model of mesial temporal lobe epilepsy. Front. Neurol. 9:632. doi : 10.3389/fneur.2018.00632
Mateos, S. S., Sánchez, C. L., Paredes, S. D., Barriga, C., and Rodríguez, A. B. (2009). Circadian levels of serotonin in plasma and brain after oral administration of tryptophan in rats. Basic Clin. Pharmacol. Toxicol. 104, 52–59. doi : 10.1111/j. 1742-7843.2008.00333.x
Mattson, R. H., Pratt, K. L., and Calverley, J. (1965). Electroencephalograms of epileptics following sleep deprivation. Arch. Neurol. 13, 310–315. doi : 10.1001/archneur.1965.00470030090009
Mcginty, D. J., and Harper, R. M. (1976). Dorsal raphe neurons : depression of firing during sleep in cats. Brain Res. 101, 569–575. doi : 10.1016/0006- 8993(76)90480-7
Mckay, L. C., and Feldman, J. L. (2008). Unilateral ablation of pre-bötzinger complex disrupts breathing during sleep but not wakefulness. Am. J. Respir. Crit. Care Med. 178, 89–95. doi : 10.1164/rccm.200712-1901OC
Meny, R., Carroll, J., Carbone, M., and Kelly, D. (1994). Cardiorespiratory recordings from infants dying suddenly and unexpectedly at home. Pediatrics 93, 44–49. doi : 10.1542/peds.93.1.44
Mohawk, J. A., Green, C. B., and Takahashi, J. S. (2012). Central and peripheral circadian clocks in mammals. Ann. Rev. Neurosci. 35, 445–462. doi : 10.1146/annurev-neuro-060909-153128
Montserrat, J. M., Kosmas, E. N., Cosio, M. G., and Kimoff, R. J. (1996). Mechanism of apnea lengthening across the night in obstructive sleep apnea. Am. J. Respir. Crit. Care Med. 154, 988–993. doi : 10.1164/ajrccm.154.4.8887596
Moore, B. M., Jerry Jou, C., Tatalovic, M., Kaufman, E. S., Kline, D. D., and Kunze, D. L. (2014). The Kv1.1 null mouse, a model of sudden unexpected death in epilepsy (SUDEP). Epilepsia 55, 1808–1816. doi : 10.1111/epi.12793
Morgan, W. W., Mcfadin, L. S., and Harvey, C. Y. (1973). A daily rhythm in norepinephrine content in regions of the hamster brain. Comp. Gen. Pharmacol. 4, 47–52. doi : 10.1016/0010-4035(73)90020-7
Mortola, J. P. (2004). Breathing around the clock : an overview of the circadian pattern of respiration. Eur. J. Appl. Physiol. 91, 119–129. doi : 10.1007/s00421-003- 0978-0
Muñoz-Ortiz, J., Muñoz-Ortiz, E., López-Meraz, L., Beltran-Parrazal, L., and Morgado-Valle, C. (2019). The pre-bötzinger complex : generation and modulation of respiratory rhythm. Neurologia (Engl Ed) 34, 461–468. doi : 10.1088/1361- 6560/ac8410
Murugesan, A., Rani, M. R. S., Vilella, L., Lacuey, N., Hampson, J. P., Faingold, C. L., et al. (2019). Postictal serotonin levels are associated with peri-ictal apnea. Neurology 93, e1485–e1494.
doi : 10.1212/WNL.00000000 00008244
Nakase, K., Kollmar, R., Lazar, J., Arjomandi, H., Sundaram, K., Silverman, J., et al. (2016). Laryngospasm, central and obstructive apnea during seizures : Defining pathophysiology for sudden death in a rat model. Epilepsy Res. 128, 126–139. doi : 10.1016/j.eplepsyres.2016.08.004
Nashef, L., Garner, S., Sander, J. W., Fish, D. R., and Shorvon, S. D. (1998). Circumstances of death in sudden death in epilepsy : interviews of bereaved relatives. J. Neurol. Neurosurg. Psychiatry 64, 349–352. doi : 10.1136/jnnp.64. 3.349
Nashef, L., Walker, F., Allen, P., Sander, J. W., Shorvon, S. D., and Fish, D. R. (1996). Apnoea and bradycardia during epileptic seizures : relation to sudden death in epilepsy. J. Neurol. Neurosurg. Psychiatry 60, 297–300. doi : 10.1136/jnnp.60.3
Nattie, E., and Li, A. (2012). “Chapter 4 - respiration and autonomic regulation and orexin,” in Progress In Brain Research (Vol. 198), ed A. Shekhar (Amsterdam, Netherlands : Elsevier), 25–46. doi : 10.1016/B978-0-444-59489-1.00004-5 Ng, M., and Pavlova, M. (2013). Why are seizures rare in rapid eye movement
sleep ? Review of the frequency of seizures in different sleep stages. Epilepsy Res. Treat. 2013:932790. doi : 10.1155/2013/932790
Nilsson, L., Farahmand, B. Y., Persson, P. G., Thiblin, I., and Tomson, T. (1999). Risk factors for sudden unexpected death in epilepsy : a case control study. Lancet 353, 888–893. doi : 10.1016/s0140-6736(98)
05114-9
Nishimura, Y., Saito, Y., Kondo, N., Matsuda, E., Fujiyama, M., Morizane, R., et al. (2015). Ictal central apnea and bradycardia in temporal lobe epilepsy complicated by obstructive sleep apnea syndrome. Epilepsy Behav. Case Rep. 4, 41–44. doi : 10.1016/j.ebcr.2015.05.001
Nobili, L., Proserpio, P., Rubboli, G., Montano, N., Didato, G., and Tassinari, C. A. (2011). Sudden unexpected death in epilepsy (SUDEP) and sleep. Sleep Med. Rev. 15, 237–246. doi : 10.1016/j.smrv.2010.07.006
Nobis, W. P., Schuele, S., Templer, J. W., Zhou, G., Lane, G., Rosenow, J. M., et al. (2018). Amygdala-stimulation-induced apnea is attention and nasal-breathing dependent. Ann. Neurol. 83, 460–471. doi : 10.1002/ana.25178
Opeskin, K., and Berkovic, S. F. (2003). Risk factors for sudden unexpected death in epilepsy : a controlled prospective study based on coroners cases. Seizure 12, 456–464. doi : 10.1016/s1059-1311(02)00352-7
Oudiette, D., Dodet, P., Ledard, N., Artru, E., Rachidi, I., Similowski, T., et al. (2018). REM sleep respiratory behaviours match mental content in narcoleptic
lucid dreamers. Sci. Rep. 8:6128. doi : 10.1038/s41598-018-24179-4
Peever, J. H., and Stephenson, R. (1997). Day-night differences in the respiratory
response to hypercapnia in awake adult rats. Respir. Physiol. 109, 241–248.
doi : 10.1016/s0034-5687(97)00056-x
Peng, W., Danison, J. L., and Seyal, M. (2017). Postictal generalized EEG
suppression and respiratory dysfunction following generalized tonic-clonic
seizures in sleep and wakefulness. Epilepsia 58, 1409–1414. doi : 10.1111/epi.13805
Petrucci, A. N., Joyal, K. G., Purnell, B. S., and Buchanan, G. F. (2020). Serotonin
and sudden unexpected death in epilepsy. Exp. Neurol. 325:113145. doi : 10.1016/j.
expneurol.2019.113145
Phabphal, K., Koonalintip, P., Sithinamsuwan, P., Wongsritrang, K.,
Amornpojnimman, T., Ekpitakdamrong, N., et al. (2021). Obstructive sleep apnea
and sudden unexpected death in epilepsy in unselected patients with epilepsy :
are they associated ? Sleep Breath. 25, 1919–1924. doi : 10.1007/s11325-021
– 02307-1
Popkirov, S., Stone, J., and Derry, C. P. (2019). Abnormal sleep in patients with
epileptic or dissociative (non-epileptic) seizures : a polysomnography study. Eur.
J. Neurol. 26, 255–260. doi : 10.1111/ene.13798
Porkka-Heiskanen, T., Strecker, R. E., and Mccarley, R. W. (2000). Brain
site-specificity of extracellular adenosine concentration changes during sleep
deprivation and spontaneous sleep : an in vivo microdialysis study. Neuroscience
99, 507–517. doi : 10.1016/s0306-4522(00)00220-7
Pramsohler, S., Schilz, R., Patzak, A., Rausch, L., and Netzer, N. C.
(2019). Periodic breathing in healthy young adults in normobaric hypoxia
equivalent to 3500 m, 4500 m and 5500 m altitude. Sleep Breath. 23, 703–709.
doi : 10.1007/s11325-019-01829-z
Pratt, K. L., Mattson, R. H., Weikers, N. J., and Williams, R. (1968). EEG
activation of epileptics following sleep deprivation : a prospective study of
114 cases. Electroencephalogr. Clin. Neurophysiol. 24, 11–15. doi : 10.1016/0013-
4694(68)90061-8
Ptak, K., Yamanishi, T., Aungst, J., Milescu, L. S., Zhang, R., Richerson, G. B.,
et al. (2009). Raphé neurons stimulate respiratory circuit activity by multiple
mechanisms via endogenously released serotonin and substance P. J. Neurosci. 29,
3720–3737. doi : 10.1523/JNEUROSCI.5271-08.2009
Purnell, B. S., and Buchanan, G. F. (2020). Free-running circadian breathing
rhythms are eliminated by suprachiasmatic nucleus lesion. J. Appl. Physiol. (1985)
129, 49–57. doi : 10.1152/japplphysiol.00211.2020
Purnell, B. S., Hajek, M. A., and Buchanan, G. F. (2017). Time-of-day influences
on respiratory sequelae following maximal electroshock-induced seizures in mice.
J Neurophysiol. 118, 2592–2600. doi : 10.1152/jn.00039.2017
Purnell, B. S., Murugan, M., Jani, R., and Boison, D. (2021a). The
good, the bad and the deadly : adenosinergic mechanisms underlying sudden
unexpected death in epilepsy. Front. Neurosci. 15:708304. doi : 10.3389/fnins.2021.
708304
Purnell, B. S., Petrucci, A. N., Li, R., and Buchanan, G. F. (2021b). The effect of
time-of-day and circadian phase on vulnerability to seizure-induced death in two
mouse models. J. Physiol. 599, 1885–1899. doi : 10.1113/JP280856
Purnell, B. S., Thijs, R. D., and Buchanan, G. F. (2018). Dead in the night :
sleep-wake and time-of-day influences on sudden unexpected death in epilepsy.
Front. Neurol. 9:1079. doi : 10.3389/fneur.2018.01079
Rao, M. L., Gross, G., Strebel, B., Halaris, A., Huber, G., Bräunig, P., et al. (1994).
Circadian rhythm of tryptophan, serotonin, melatonin and pituitary hormones
in schizophrenia. Biol. Psychiatry 35, 151–163. doi : 10.1016/0006-3223(94)
91147-9
Re, C. J., Batterman, A. I., Gerstner, J. R., Buono, R. J., and Ferraro, T. N. (2020).
The molecular genetic interaction between circadian rhythms and susceptibility to
seizures and epilepsy. Front. Neurol. 11:520. doi : 10.3389/fneur.2020.00520
Reed, D. J., and Kellogg, R. H. (1958). Changes in respiratory response to CO2
during natural sleep at sea level and at altitude. J. Appl. Physiol. 13, 325–330.
doi : 10.1152/jappl.1958.13.3.325
Remmers, J. E., Degroot, W. J., Sauerland, E. K., and Anch, A. M. (1978).
Pathogenesis of upper airway occlusion during sleep. J. Appl. Physiol. Respir.
Environ. Exerc. Physiol. 44, 931–938. doi : 10.1152/jappl.1978.44.6.931
Rhone, A. E., Kovach, C. K., Harmata, G. I., Sullivan, A. W., Tranel, D.,
Ciliberto, M. A., et al. (2020). A human amygdala site that inhibits respiration and
elicits apnea in pediatric epilepsy. JCI Insight 5:e134852. doi : 10.1172/jci.insight.
134852
Richerson, G. B. (1995). Response to CO2 of neurons in the rostral ventral
medulla in vitro. J. Neurophysiol. 73, 933–944. doi : 10.1152/jn.1995.73.3.933
Richerson, G. B. (2004). Serotonergic neurons as carbon dioxide sensors that
maintain pH homeostasis. Nat. Rev. Neurosci. 5, 449–461. doi : 10.1038/nrn1409
Richerson, G. B. (2013). Serotonin : the anti-suddendeathamine ? Epilepsy Curr.
13, 241–244. doi : 10.5698/1535-7597-13.5.241
Richerson, G. B., and Buchanan, G. F. (2011). The serotonin axis : shared
mechanisms in seizures, depression and SUDEP. Epilepsia 52, 28–38.
doi : 10.1111/j.1528-1167.2010.02908.x
Rosenwasser, A. M., Trubowitsch, G., and Adler, N. T. (1985). Circadian rhythm
in metabolic activity of suprachiasmatic, supraoptic and raphe nuclei. Neurosci.
Lett. 58, 183–187. doi : 10.1016/0304-3940(85)90161-2
Roundtree, H. M., Simeone, T. A., Johnson, C., Matthews, S. A., Samson, K. K.,
and Simeone, K. A. (2016). Orexin receptor antagonism improves sleep and
reduces seizures in kcna1-null mice. Sleep 39, 357–368. doi : 10.5665/sleep.5444
Rugg-Gunn, F., Duncan, J., Hjalgrim, H., Seyal, M., and Bateman, L. (2016).
From unwitnessed fatality to witnessed rescue : nonpharmacologic interventions
in sudden unexpected death in epilepsy. Epilepsia 57, 26–34. doi : 10.1111/epi.
13231
Ryvlin, P., Nashef, L., Lhatoo, S. D., Bateman, L. M., Bird, J., Bleasel, A.,
et al. (2013). Incidence and mechanisms of cardiorespiratory arrests in epilepsy
monitoring units (MORTEMUS) : a retrospective study. Lancet Neurol. 12,
966–977. doi : 10.1016/S1474-4422(13)70214-X
Ryvlin, P., Rheims, S., and Risse, G. (2006). Nocturnal frontal lobe epilepsy.
Epilepsia 47, 83–86. doi : 10.1111/j.1528-1167.2006.00698.x
Ryvlin, P., So, E. L., Gordon, C. M., Hesdorffer, D. C., Sperling, M. R.,
Devinsky, O., et al. (2018). Long-term surveillance of SUDEP in drug-resistant
epilepsy patients treated with VNS therapy. Epilepsia 59, 562–572. doi : 10.1111/epi.
14002
14002
Saaresranta, T., Polo-Kantola, P. I., Irjala, K., Helenius, H., and Polo, O. (1999).
Respiratory insufficiency in postmenopausal women : sustained improvement
of gas exchange with short-term medroxyprogesterone acetate. Chest 115,
1581–1587. doi : 10.1378/chest.115.6.1581
Sainju, R. K., Dragon, D. N., Winnike, H. B., Nashelsky, M. B., Granner, M. A.,
Gehlbach, B. K., et al. (2019). Ventilatory response to CO2 in patients with epilepsy.
Epilepsia 60, 508–517. doi : 10.1111/epi.14660
Sainju, R. K., Dragon, D. N., Winnike, H. B., Ten Eyck, P., Granner, M. A.,
Gehlbach, B. K., et al. (2021). Hypercapnic ventilatory response in epilepsy patients
treated with VNS : a case-control study. Epilepsia 62, e140–e146. doi : 10.1111/epi.
16997
Sakai, K. (2011). Sleep-waking discharge profiles of dorsal raphe nucleus neurons
in mice. Neuroscience 197, 200–224. doi : 10.1016/j.neuroscience.2011.09.024
Sakai, K., and Crochet, S. (2001). Differentiation of presumed serotonergic dorsal
raphe neurons in relation to behavior and wake-sleep states. Neuroscience 104,
1141–1155. doi : 10.1016/s0306-4522(01)00103-8
Sakurai, T. (2007). The neural circuit of orexin (hypocretin) : maintaining sleep
and wakefulness. Nat. Rev. Neurosci. 8, 171–181. doi : 10.1038/nrn2092
Sanchez, R. E. A., Bussi, I. L., Ben-Hamo, M., Caldart, C. S., Catterall, W. A.,
and De La Iglesia, H. O. (2019). Circadian regulation of sleep in a pre-clinical
model of dravet syndrome : dynamics of sleep stage and siesta re-entrainment. Sleep
42:zsz173. doi : 10.1093/sleep/zsz173
Santos, E. A. d. S., Marques, T. E. B. S., Matos, H. D. C., Leite, J. P., GarciaCairasco, N., Paçó-Larson, M. L., et al. (2015). Diurnal variation has effect on
differential gene expression analysis in the hippocampus of the pilocarpineinduced model of mesial temporal lobe epilepsy. PLoS One 10:e0141121.
doi : 10.1371/journal.pone.0141121
Saper, C. B., Scammell, T. E., and Lu, J. (2005). Hypothalamic regulation of sleep
and circadian rhythms. Nature 437, 1257–1263. doi : 10.1038/nature04284
Scharf, M. T., Greenberg, P., Wong, S., and Mani, R. (2020). Obstructive sleep
apnea risk in patients with focal versus generalized epilepsy. Epilepsy Behav.
111:107190. doi : 10.1016/j.yebeh.2020.107190
Seifert, E. L., Knowles, J., and Mortola, J. P. (2000). Continuous circadian
measurements of ventilation in behaving adult rats. Respir. Physiol. 120, 179–183.
doi : 10.1016/s0034-5687(00)00108-0
Seifert, E. L., and Mortola, J. P. (2002). The circadian pattern of breathing
in conscious adult rats. Respir. Physiol. 129, 297–305. doi : 10.1016/s0034-
5687(01)00316-4
Semmelroch, M., Elwes, R. D., Lozsadi, D. A., and Nashef, L. (2012).
Retrospective audit of postictal generalized EEG suppression in telemetry.
Epilepsia 53, e21–e24. doi : 10.1111/j.1528-1167.2011.03296.x
Severson, C. A., Wang, W., Pieribone, V. A., Dohle, C. I., and Richerson, G. B.
(2003). Midbrain serotonergic neurons are central pH chemoreceptors. Nat.
Neurosci. 6, 1139–1140. doi : 10.1038/nn1130
Seyal, M., Bateman, L. M., Albertson, T. E., Lin, T.-C., and Li, C.-S. (2010).
Respiratory changes with seizures in localization-related epilepsy : analysis of
periictal hypercapnia and airflow patterns. Epilepsia 51, 1359–1364. doi : 10.1111/j.
1528-1167.2009.02518.x
Seyal, M., Pascual, F., Lee, C. Y., Li, C. S., and Bateman, L. M. (2011). Seizurerelated cardiac repolarization abnormalities are associated with ictal hypoxemia.
Epilepsia 52, 2105–2111. doi : 10.1111/j.1528-1167.2011.03262.x
Shankar, R., Cox, D., Jalihal, V., Brown, S., Hanna, J., and Mclean, B. (2013).
Sudden unexpected death in epilepsy (SUDEP) : development of a safety checklist.
Seizure 22, 812–817. doi : 10.1016/j.seizure.2013.07.014
Shen, H.-Y., Li, T., and Boison, D. (2010). A novel mouse model
for sudden unexpected death in epilepsy (SUDEP) : role of impaired
adenosine clearance. Epilepsia 51, 465–468. doi : 10.1111/j.1528-1167.2009.
02248.x
Shouse, M. N. (1988). Sleep deprivation increases susceptibility to kindled and
penicillin seizure events during all waking and sleep states in cats. Sleep 11,
162–171. doi : 10.1093/sleep/11.2.162
Shouse, M. N., and Sterman, M. B. (1982). Acute sleep deprivation
reduces amygdala-kindled seizure thresholds in cats. Exp. Neurol. 78, 716–727.
doi : 10.1016/0014-4886(82)90086-3
Siekierka, M., Tafil-Klawe, M., Adamczyk, W., Klawe, J. J., and Złomanczuk, P. ´
(2007). Low amplitude daily changes in reflex ventilatory response to progressive
isocapnic hypoxia. J. Physiol. Pharmacol. 58, 633–637.
Simeone, K. A., Hallgren, J., Bockman, C. S., Aggarwal, A., Kansal, V.,
Netzel, L., et al. (2018). Respiratory dysfunction progresses with age in kcna1-null
mice, a model of sudden unexpected death in epilepsy. Epilepsia 59, 345–357.
doi : 10.1111/epi.13971
Simonds, A. K. (1994). Sleep studies of respiratory function and home
respiratory support. BMJ 309, 35–40. doi : 10.1136/bmj.309.6946.35
Smith, J. C., Ellenberger, H. H., Ballanyi, K., Richter, D. W., and Feldman, J. L.
(1991). Pre-bötzinger complex : a brainstem region that may generate respiratory
rhythm in mammals. Science 254, 726–729. doi : 10.1126/science.1683005
Smith, H. R., Leibold, N. K., Rappoport, D. A., Ginapp, C. M., Purnell, B. S.,
Bode, N. M., et al. (2018). Dorsal raphe serotonin neurons mediate CO2-induced
arousal from sleep. J. Neurosci. 38, 1915–1925. doi : 10.1523/JNEUROSCI.2182-17.
2018
Snyder, F., Hobson, J. A., Morrison, D. F., and Goldfrank, F. (1964). Changes in
respiration, heart rate and systolic blood pressure in human sleep. J. Appl. Physiol.
19, 417–422. doi : 10.1152/jappl.1964.19.3.417
Somboon, T., Grigg-Damberger, M. M., and Foldvary-Schaefer, N. (2019).
Epilepsy and sleep-related breathing disturbances. Chest 156, 172–181.
doi : 10.1016/j.chest.2019.01.016
Sowers, L. P., Massey, C. A., Gehlbach, B. K., Granner, M. A., and
Richerson, G. B. (2013). Sudden unexpected death in epilepsy : fatal post-ictal
respiratory and arousal mechanisms. Respir. Physiol. Neurobiol. 189, 315–323.
doi : 10.1016/j.resp.2013.05.010
Spengler, C. M., Czeisler, C. A., and Shea, S. A. (2000). An endogenous circadian
rhythm of respiratory control in humans. J. Physiol. 526, 683–694. doi : 10.1111/j.
1469-7793.2000.00683.x
Spengler, C. M., and Shea, S. A. (2000). Endogenous circadian rhythm of
pulmonary function in healthy humans. Am. J. Respir. Crit. Care Med. 162,
1038–1046. doi : 10.1164/ajrccm.162.3.9911107
Sridhar, R., Thach, B. T., Kelly, D. H., and Henslee, J. A. (2003). Characterization
of successful and failed autoresuscitation in human infants, including those dying
of SIDS. Pediatr. Pulmonol. 36, 113–122. doi : 10.1002/ppul.10287
Stephenson, R., Liao, K. S., Hamrahi, H., and Horner, R. L. (2001). Circadian
rhythms and sleep have additive effects on respiration in the rat. J. Physiol. 536,
225–235. doi : 10.1111/j.1469-7793.2001.00225.x
Stewart, M. (2018). An explanation for sudden death in epilepsy (SUDEP).
J. Physiol. Sci. 68, 307–320. doi : 10.1007/s12576-018-0602-z
Stöllberger, C., and Finsterer, J. (2004). Cardiorespiratory findings in sudden
unexplained/unexpected death in epilepsy (SUDEP). Epilepsy Res. 59, 51–60.
doi : 10.1016/j.eplepsyres.2004.03.008
Stupfel, M., Damiani, P., Perramon, A., Busnel, M. C., Gourlet, V., and
Thierry, H. (1985). Ultradian and circadian respiratory rhythms in grouped
small laboratory vertebrate species as a method to assess the effects of
environmental challenges. Comp. Biochem. Physiol. Comp. A Physiol. 80, 225–231.
doi : 10.1016/0300-9629(85)90545-6
Stupfel, M., and Pletan, Y. (1983). Respiratory ultradian rhythms of mean
and low frequencies : a comparative physiological approach. Chronobiologia 10,
283–292.
Surges, R., and Sander, J. W. (2012). Sudden unexpected death in epilepsy :
mechanisms, prevalence and prevention. Curr. Opin. Neurol. 25, 201–207.
doi : 10.1097/WCO.0b013e3283506714
Surges, R., Thijs, R. D., Tan, H. L., and Sander, J. W. (2009). Sudden unexpected
death in epilepsy : risk factors and potential pathomechanisms. Nat. Rev. Neurol. 5,
492–504. doi : 10.1038/nrneurol.2009.118
Sveinsson, O., Andersson, T., Carlsson, S., and Tomson, T. (2018). Circumstances
of SUDEP : a nationwide population-based case series. Epilepsia 59, 1074–1082.
doi : 10.1111/epi.14079
Sveinsson, O., Andersson, T., Mattsson, P., Carlsson, S., and Tomson, T. (2020).
Clinical risk factors in SUDEP : a nationwide population-based case-control study.
Neurology 94, e419–e429. doi : 10.1212/WNL.0000000000008741
Tanaka, T., and Naquet, R. (1975). Kindling effect and sleep organization
in cats. Electroencephalogr. Clin. Neurophysiol. 39, 449–454. doi : 10.1016/0013-
4694(75)90045-0
Tao, J. X., Qian, S., Baldwin, M., Chen, X. J., Rose, S., Ebersole, S. H., et al. (2010).
SUDEP, suspected positional airway obstruction and hypoventilation in postictal
coma. Epilepsia 51, 2344–2347. doi : 10.1111/j.1528-1167.2010.02719.x
Tao, J. X., Sandra, R., Wu, S., and Ebersole, J. S. (2015). Should the “back to
sleep” campaign be advocated for SUDEP prevention ? Epilepsy Behav. 45, 79–80.
doi : 10.1016/j.yebeh.2015.02.020
Tennis, P., Cole, T. B., Annegers, J. F., Leestma, J. E., Mcnutt, M., and Rajput, A.
(1995). Cohort study of incidence of sudden unexplained death in persons
with seizure disorder treated with antiepileptic drugs in Saskatchewan, Canada.
Epilepsia 36, 29–36. doi : 10.1111/j.1528-1157.1995.tb01661.x
Teran, F. A., Kim, Y., Crotts, M. S., Bravo, E., Emaus, K. J., Richerson, G. B.
(2019). Time of day and a ketogenic diet influence susceptibility to SUDEP in
Scn1aR1407X/+ mice. Front. Neurol. 10:278. doi : 10.3389/fneur.2019.00278
Tio, E., Culler, G. W., Bachman, E. M., and Schuele, S. (2020). Ictal central apneas
in temporal lobe epilepsies. Epilepsy Behav. 112:107434. doi : 10.1016/j.yebeh.2020.
107434
Tochikubo, O., Ikeda, A., Miyajima, E., and Ishii, M. (1996). Effects of
insufficient sleep on blood pressure monitored by a new multibiomedical recorder.
Hypertension 27, 1318–1324. doi : 10.1161/01.hyp.27.6.1318
Totola, L. T., Malheiros-Lima, M. R., Delfino-Pereira, P., Del Vecchio, F.,
Souza, F. C., Takakura, A. C., et al. (2019). Amygdala rapid kindling impairs
breathing in response to chemoreflex activation. Brain Res. 1718, 159–168.
doi : 10.1016/j.brainres.2019.05.015
Tupal, S., and Faingold, C. L. (2006). Evidence supporting a role of serotonin
in modulation of sudden death induced by seizures in DBA/2 mice. Epilepsia 47,
21–26. doi : 10.1111/j.1528-1167.2006.00365.x
Tupal, S., and Faingold, C. L. (2019). Fenfluramine, a serotonin-releasing
drug, prevents seizure-induced respiratory arrest and is anticonvulsant in the
DBA/1 mouse model of SUDEP. Epilepsia 60, 485–494. doi : 10.1111/epi.
14658
Tusiewicz, K., Moldofsky, H., Bryan, A. C., and Bryan, M. H. (1977).
Mechanics of the rib cage and diaphragm during sleep. J. Appl. Physiol.
Respir. Environ. Exerc. Physiol. 43, 600–602. doi : 10.1152/jappl.1977.
43.4.600
Van Der Lende, M., Hesdorffer, D., Sander, J., and Thijs, R. (2018). Nocturnal
supervision and SUDEP risk at different epilepsy care settings. Neurology 91,
e1508–e1518. doi : 10.1212/WNL.0000000000006356
Van Gompel, J. J., Bower, M. R., Worrell, G. A., Stead, M., Chang, S.-Y.,
Goerss, S. J., et al. (2014). Increased cortical extracellular adenosine correlates with
seizure termination. Epilepsia 55, 233–244. doi : 10.1111/epi.12511
Vaughn, B. V., and D’cruz, O. F. (2004). Sleep and epilepsy. Semin. Neurol. 24,
301–313. doi : 10.1055/s-2004-835068
Vendrame, M., Jackson, S., Syed, S., Kothare, S. V., and Auerbach, S. H. (2014).
Central sleep apnea and complex sleep apnea in patients with epilepsy. Sleep Breath.
18, 119–124. doi : 10.1007/s11325-013-0858-8
Venit, E. L., Shepard, B. D., and Seyfried, T. N. (2004). Oxygenation prevents
sudden death in seizure-prone mice. Epilepsia 45, 993–996. doi : 10.1111/j.0013-
9580.2004.02304.x
Vilella, L., Lacuey, N., Hampson, J. P., Rani, M. R. S., Sainju, R. K., Friedman, D.,
et al. (2019). Postconvulsive central apnea as a biomarker for sudden unexpected
death in epilepsy (SUDEP). Neurology 92, e171–e182. doi : 10.1212/WNL.
0000000000006785
Wang, W., Pizzonia, J. H., and Richerson, G. B. (1998). Chemosensitivity of
rat medullary raphe neurones in primary tissue culture. J. Physiol. 511, 433–450.
doi : 10.1111/j.1469-7793.1998.433bh.x
Wessberg, P., Hedner, J., Hedner, T., Persson, B., and Jonason, J. (1984).
Adenosine mechanisms in the regulation of breathing in the rat. Eur. J. Pharmacol.
106, 59–67. doi : 10.1016/0014-2999(84)90678-2
Wu, H., Liu, Y., Liu, L., Meng, Q., Du, C., Li, K., et al. (2021). Decreased
expression of the clock gene bmal1 is involved in the pathogenesis of temporal
lobe epilepsy. Mol. Brain 14:113. doi : 10.1186/s13041-021-00824-4
Yoshida, Y., Fujiki, N., Nakajima, T., Ripley, B., Matsumura, H., Yoneda, H.,
et al. (2001). Fluctuation of extracellular hypocretin-1 (orexin A) levels in the rat
in relation to the light-dark cycle and sleep-wake activities. Eur. J. Neurosci. 14,
1075–1081. doi : 10.1046/j.0953-816x.2001.01725.x
Young, T., Palta, M., Dempsey, J., Skatrud, J., Weber, S., and Badr, S. (1993). The occurrence of sleep-disordered breathing among middle-aged adults. N. Engl. J. Med. 328, 1230–1235.
doi : 10.1056/NEJM199304293 281704
Yuan, H., Pinto, S. J., Huang, J., Mcdonough, J. M., Ward, M. B., Lee, Y. N., et al. (2012). Ventilatory responses to hypercapnia during wakefulness and sleep in obese adolescents with and without obstructive sleep apnea syndrome. Sleep 35, 1257–1267. doi : 10.5665/sleep.2082
Zambrelli, E., Saibene, A. M., Furia, F., Chiesa, V., Vignoli, A., Pipolo, C., et al. (2016). Laryngeal motility alteration : a missing link between sleep apnea and vagus nerve stimulation for epilepsy. Epilepsia 57, e24–e27. doi : 10.1111/epi. 13252
Zeng, C., Long, X., Cotten, J. F., Forman, S. A., Solt, K., Faingold, C. L., et al. (2015). Fluoxetine prevents respiratory arrest without enhancing ventilation in DBA/1 mice. Epilepsy Behav. 45, 1–7. doi : 10.1016/j.yebeh.2015.02.013
Zhan, Q., Buchanan, G. F., Motelow, J. E., Andrews, J., Vitkovs.kiy, P., Chen, W. C., et al. (2016). Impaired serotonergic brainstem function during and after seizures. J. Neurosci. 36, 2711–2722. doi : 10.1523/JNEUROSCI.4331-15.2016
Zhang, R., Tan, Z., Niu, J., and Feng, H.-J. (2021). Adrenergic α2 receptors are implicated in seizure-induced respiratory arrest in DBA/1 mice. Life Sci. 284:119912. doi : 10.1016/j.lfs.2021.119912
Zhang, H., Zhao, H., and Feng, H. J. (2017). Atomoxetine, a norepinephrine reuptake inhibitor, reduces seizure-induced respiratory arrest. Epilepsy Behav. 73, 6–9. doi : 10.1016/j.yebeh.2017.04.046
Zhao, H., Cotten, J. F., Long, X., and Feng, H. J. (2017). The effect of atomoxetine, a selective norepinephrine reuptake inhibitor, on respiratory arrest and cardiorespiratory function in the DBA/1 mouse model of SUDEP. Epilepsy Res. 137, 139–144. doi : 10.1016/j.eplepsyres.2017.08.005
Zhuo, L., Zhang, Y., Zielke, H. R., Levine, B., Zhang, X., Chang, L., et al. (2012). Sudden unexpected death in epilepsy : evaluation of forensic autopsy cases. Forensic Sci. Int. 223, 171–175. doi : 10.1016/j.forsciint.2012.08.02
Financement
Ce travail publié a été soutenu par les subventions F31NS125955 (à KJ), R01NS095842 (à GB) des National Institutes of Health/National Institute of Neurological Disorders and Stroke, la chaire Beth L. Tross d’épilepsie du Carver College of Medicine de l’Université de l’Iowa (à GB) et la subvention T32GM0073367 (à BK) des National Institutes of Health/National Institute of General Medicine Sciences.
Déclarations
Contributions des auteurs
KJ et BK ont rédigé la première version du document. Le manuscrit final a été révisé et approuvé par KJ, BK et GB. Tous les auteurs ont contribué à l’article et approuvé la version soumise.
Conflit d’intérêts
Les auteurs déclarent que la recherche a été menée en l’absence de toute relation commerciale ou financière pouvant être interprétée comme un conflit d’intérêts potentiel.
Note de l’éditeur
Les opinions exprimées dans cet article n’engagent que leurs auteurs et ne reflètent pas nécessairement celles de leurs organisations affiliées, ni celles de l’éditeur, des rédacteurs ou des relecteurs. L’éditeur n’offre aucune garantie ni n’approuve aucun produit évalué dans cet article, ni aucune allégation formulée par son fabricant.